
Гіп про фіз (від грец. Hyp ó physis - відросток), нижній мозковий придаток (hypophysis cerebri, glandula pituitaria), заліза з внутрішньою секрецією , Яка грає у всіх хребетних тварин і у людини провідну роль в гормональної регуляції . Г. розташований в турецькому сідлі основної кістки черепа, біля основи головного мозку і пов'язаний з ним за допомогою ніжки (воронки), що представляє собою виріст дна 3-го мозкового шлуночка. Форма, розмір і вага Г. різні в різних видів і залежать від віку та фізіологічного стану організму. У людини Г. важить 0,5-0,6 г. У Г. розрізняють три долі: передню (залізисту), середню (проміжну) і задню (нервову). Передня і середня частки закладаються у зародка у вигляді випинання епітелію даху первинної ротової порожнини; задня частка утворюється з дна воронки проміжного мозку. Ембріональний зачаток передньої і середньої часток в подальшому відділяється від епітелію первинної ротової порожнини, зростає у напрямку до мозку і зростається з зачатком задньої долі. Лише у деяких хрящових риб зв'язок передньої долі Р. з епітелієм первинної ротової порожнини зберігається і у дорослих організмів. У одних ссавців, наприклад у кішки, задня доля Р. має порожнину, що сполучається з порожниною 3-го шлуночка, у інших, наприклад у собаки, порожнина зберігається лише в ніжці, що сполучає Г. з проміжним мозком; у деяких ссавців (наприклад, у кролика і у всіх приматів) задня частка і ніжка Г. позбавлені порожнини і є щільними освіти. У дорослого організму Р. тісно пов'язаний анатомічно з головним мозком. Г. забезпечений великою кількістю нервових волокон, що вступають в нього через ніжку з гіпоталамічної області (див. гіпоталамус ) І по стінках гіпофізарних артерій - з нервового сонного сплетення.
Передня доля Р. дорослого організму складається з залозистого епітелію, в якому виділяють 3 типи клітин, що розрізняються за здатністю забарвлюватися кислими або основними фарбами: хромофобние, або головні, клітки; оксифільні, або еозинофільні, клітини і базофільні клітини. Хромофобние клітини - резервний матеріал, з якого розвиваються оксифільні і базофільні клітини. Співвідношення оксифільних і базофільних клітин в передній долі Г. змінюється в залежності від статі, віку і фізіологічного стану організму. Так, після видалення щитовидної залози (тиреоїдектомія) кількість оксифільних клітин різко зменшується, аж до повного зникнення, базофільні клітини, дегенеріруя, перетворюються в т. Н. клітини тиреоїдектомії; після кастрації базофільні клітини гіпертрофуються і перетворюються в т. н. клітини кастрації. Зміни в клітинному складі передньої долі Р., що настають після видалення щитовидної залози або кастрації, можуть запобігти або усунуті введенням тироксину або статевих гормонів . Середня частка Г. складається з епітеліальної тканини. Задня доля Р. утворена нейроглії , В якій містяться великі пірамідальні або веретеноподібні клітини, т. Н. пітуіціти. Найбільш складна і різноманітна фізіологічна роль передньої долі Р., від нормальної функції якої залежать зростання і розмноження, основний, вуглеводний, мінеральний, жировий і білковий обмін. З екстракту передньої долі Р. виділено 7 гормонів: гормон росту, або соматотропний гормон, тиреотропний гормон, фолікулостимулюючий гормон, лютеїнізуючий гормон, лютеотропного гормон, пролактин (лактогенний) і адренокортикотропний гормон (АКТГ). Всі гормони передньої долі мають білкову природу і отримані в очищеному вигляді, деякі з них, наприклад гормон росту і лактогенний, виділені в кристалічній формі, ін. Синтезовані (наприклад, АКТГ). тиреотропний і гонадотропні гормони продукуються базофільними клітинами, які відповідно до цього ділять на два типи: т. н. тіреотрофов і гонадотрофов. Оксифільні клітини виробляють гормон росту і пролактин. Питання про клітинах, які продукують АКТГ, не вирішене; ймовірно, він утворюється базофілами.
Гормон росту. Хірургічне видалення Г. (гіпофізектомія) у молодої тварини приводить до зупинки росту. Ін'єкції такою твариною гипофизарного екстракту, що містить гормон росту, відновлюють у них нормальне зростання. Введення гормону росту молодим зростаючим тваринам різко стимулює зростання і приводить до гігантизму (в експерименті були отримані гіганти амбістоми, щури, собаки і ін. Тварин): у людини надлишкове виділення гормону росту викликає захворювання з явищами гігантизму або акромегалії . Знижений виділення гормону росту обумовлює карликовий ріст (см. нанізм ). Фолликулостимулирующий, лютеїнізуючий і лютеотропного гормони. Атрофія статевої системи, що наступає після видалення Г., може бути відвернена введенням гонадотропних гормонів. У інфантильних тварин введення цих гормонів викликає передчасне статеве дозрівання. Ін'єкція гипофизарного екстракту, що містить гонадотропні гормони, жабам викликає у них ікрометання і сперматогенез в осінній і зимовий час; з ікри після запліднення розвиваються нормальні пуголовки. Фолікулостимулюючий гормон регулює зростання фолікулів в яєчниках і сперматогенез. Лютеїнізуючого гормону викликає у самок передчасний зростання фолікулів , овуляцію , Утворення жовтого тіла, а у самців - секрецію чоловічого статевого гормону проміжними клітками насінники, т. Е. Клітинами Лейдіга. Лютеотропний гормон підтримує функцію жовтого тіла; у деяких тварин (щур, вівця) цей гормон викликає лактацію. Пролактин (лактогенний гормон). Бере участь в регуляції процесу виділення молока. Видалення передньої долі Р. у лактуючих самок припиняє секрецію молока; введення пролактину відновлює лактацію. Тиреотропний гормон. Видалення передньої долі Р. викликає атрофію щитовидної залози і, як наслідок цього, зниження основного обміну. Ін'єкції гипофизарного екстракту, що містить тиреотропний гормон, викликають збільшення щитовидної залози і посилення її функції. А КТГ стимулює діяльність кори надниркових залоз і виділення нею кортикостероїдних гормонів, а також відновлює атрофовану в результаті видалення Г. залозу. Вплив передньої долі Р. на обмін речовин здійснюється через гормон росту, АКТГ і ін. Гормони.
Середня частка Г. виробляє гормон інтермедії, або меланоцитостімулірующий гормон, що впливає на забарвлення шкіри риб і земноводних. Фізіологічне значення цього гормону у птахів і ссавців неясно.
Задня доля Р. бере участь в регуляції рівня кров'яного тиску, сечовиділення (гормон вазопресин) і діяльності мускулатури матки (гормон окситоцин). Вазопресин і окситоцин утворюються в паравентрікулярних і супраоптичних ядрах гіпоталамуса, звідки вони надходять в задню частку Г. Обидва гормону синтезовані.
Функції Г. залежать від умов зовнішнього середовища. З дослідів, проведених на птахах і ссавців, встановлено, що світло регулює гонадотропну, тиреотропну і адренокортикотропну функції Г .; дію світла на Г. здійснюється через центральну нервову систему. Доведено також, що ендокринні функції Р. знаходяться під контролем гіпоталамуса, в якому виробляються особливі нейрогуморальні речовини пептидної природи - т. Зв. вивільняють, або релізінг-фактори, що стимулюють гуморальним шляхом секрецію гормонів Р. (див. Нейросекреція ).
Патологія Г. Порушення нормальної діяльності Р. можуть виражатися підвищенням ( гіперпітуітарізм ) Або ослабленням ( гіпопітуїтаризм ) Його окремих функцій, рідше - в повному їх випаданні. Підвищення внутрішньої секреції Р. виявляється розладами зростання і розвитку в дитячому віці - гігантизм, у дорослих - акромегалией. Ослаблення або випадання функцій Г. в дитячому віці призводить до затримки росту (карликовий зростання), психічного розвитку, інфантилізму , атрофії щитовидної залози і кори наднирників , Глибоких змін вуглеводного і жирового обміну, зниженню окислювальних процесів і ін .; у дорослих - до ожиріння , Припинення статевого циклу, атрофії щитовидної, статевих залоз і кори надниркових залоз та ін. В механізмі розвитку ряду т. Н. гіпофізарних захворювань ( Іценко - Кушинга хвороба , діабет нецукровий , Передчасне статеве дозрівання і ін.) Вирішальне значення мають первинні порушення діяльності гіпоталамуса.
Літ .: Киршенблат Я. Д., Загальна ендокринологія, М., 1965; Гіпоталамічна регуляція передньої частини гіпофіза, пров. з англ., Будапешт, 1965; Лейтес С. М., Лаптєва Н. Н., Нариси по патофізіології обміну речовин і ендокринної системи, М., 1967; Еськин І. А., Основи фізіології ендокринних залоз, М., 1968; Тонких А. В., гіпоталамо-гіпофізарно область і регуляція фізіологічних функцій організму, М.-Л., 1965; Schreiber V., The hypothalamo-hypophysial system, Prague, 1963.
І. А. Ескін, Л. М. Гольбер.
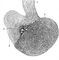
Серединний поздовжній розріз гіпофіза людини (схема): 1 - воронкоподібний відросток; 2 - задня частка; 3 - проміжна частка; 4 - передня частка.
