
Блукаю е ня, процес анаеробного розщеплення органічних речовин, переважно вуглеводів, що відбувається під впливом мікроорганізмів або виділених з них ферментів . В ході Б. в результаті пов'язаних окисно-відновних реакцій звільняється енергія, необхідна для життєдіяльності мікроорганізмів, і утворюються хімічні сполуки, які мікроорганізми використовують для біосинтезу амінокислот, білків, органічних кислот, жирів і ін. Компонентів тіла. Одночасно накопичуються кінцеві продукти Б. Залежно від їх характеру розрізняють Б. спиртове, молочнокисле, маслянокислое, пропіоновокисле, ацетоно-бутилове, ацетоно-етилове і ін. Види. Характер Б., його інтенсивність, кількісні співвідношення кінцевих продуктів, а також напрямок Б. залежать від особливостей його збудника і умов, при яких Б. протікає (pH, аерація, субстрат і ін.).
Спиртове Б. У 1836 французький учений Каньяр де ла Тур встановив, що спиртне Б. пов'язано із зростанням і розмноженням дріжджів. Хімічне рівняння спиртового Б .: C6H12O6 ® 2C2H5OH + 2CO2 було дано французькими хіміками А. Лавуазьє (тисячу сімсот вісімдесят дев'ять) і Ж. Гей-Люссак (1815). Л. Пастер прийшов до висновку (1857), що спиртове Б. можуть викликати тільки живі дріжджі в анаеробних умовах ( «бродіння - це життя без повітря»). На противагу цьому німецький учений Ю. Лібіх наполегливо наполягав на тому, що Б. відбувається поза живою клітиною. На можливість бесклеточного спиртного Б. вперше (1871) вказала російський лікар-біохімік М. М. Манассеина. Німецький хімік Е. Бухнер в 1897, віджавши під великим тиском дріжджі, розтерті з кварцовим піском, отримав безклітковий сік, зброджує цукор з утворенням спирту і CO2. При нагріванні до 50 ° C і вище сік втрачав бродильні властивості. Все це вказувало на ферментативну природу активного початку, що міститься в дріжджовому соку. Російський хімік Л. А. Іванов виявив (1905), що додані до дріжджового соку фосфати у декілька разів підвищують швидкість Б. Дослідження вітчизняних біохіміків А. І. Лебедєва, С. П. Костичева, Я. О. Парнаса і німецьких біохіміків К. Нейберга, Г. Ембдена, О. Мейергофа і ін. підтвердили, що фосфорна кислота бере участь в найважливіших етапах спиртного Б.

Надалі багато дослідників детально вивчили ферментативну природу і механізм спиртного Б. (див. Схему). Перша реакція перетворення глюкози при спиртному Б. - приєднання до глюкози під впливом ферменту глюкокинази залишку фосфорної кислоти від аденозинтрифосфорної кислоти (АТФ, див. Аденозінфосфорниє кислоти ). При цьому утворюються аденозіндіфосфорная кислота (АДФ) і глюкозо-6-фосфорна кіслотата. Остання під дією ферменту глюкозофосфаті-зомерази перетворюється в фруктозо-6-фосфорну кислоту, яка, отримуючи від нової молекули АТФ (за участю ферменту фосфофруктокинази) ще один залишок фосфорної кислоти, перетворюється в фруктозо-1,6-діфосфорная кислоту. (Ця і наступна реакції, позначені зустрічними стрілками, оборотні, тобто. Е. Їх напрямок залежить від умов - концентрації ферменту, pH і ін.) Під впливом ферменту кетозом-1-фосфатальдолази фруктозо-1,6-дифосфорная кислота розщеплюється на гліцерінальдегидфосфорной і діоксіацетонфосфорную кислоти які можуть перетворюватися один в одного під дією ферменту тріозофосфатізомерази. Гліцерінальдегидфосфорной кислота, приєднуючи молекулу неорганічної фосфорної кислоти і окислюючись під дією ферменту дегідрогенази фосфогліцерінальдегіда, активною групою якого у дріжджів є никотинамидадениндинуклеотид (НАД), перетворюється в 1,3-дифосфогліцеринової кислоту. Молекула діоксіацетонфосфорной кислоти під дією тріозофосфатізомерази дає другу молекулу гліцерінальдегидфосфорной кислоти, також піддається окисленню до 1,3-дифосфогліцеринової кислоти; остання, віддаючи АДФ (під дією ферменту фосфогліцераткинази) один залишок фосфорної кислоти, перетворюється в З-фосфогліцеріновую кислоту, яка під дією ферменту фосфогліцеро-мутази перетворюється в 2-фосфогліцеріновую кислоту, а вона під впливом ферменту фосфопіруват-гідратази - в фосфоенол-пировиноградную кислоту . Остання за участю ферменту піруваткінази передає залишок фосфорної кислоти молекулі АДФ, в результаті чого утворюється молекула АТФ і молекула енолпіровіноградной кислоти, яка дуже нестійка і переходить в пировиноградную кислоту. Ця кислота за участю наявного в дріжджах ферменту піруватдекарбоксилази розщеплюється на оцтовий альдегід і двоокис вуглецю. Оцтовий альдегід, реагуючи з утворилася при окисленні гліцерінальдегидфосфорной кислоти відновленою формою никотинамидадениндинуклеотида (НАД-Н), за участю ферменту алкогольдегідрогенази перетворюється в етиловий спирт. Сумарно рівняння спиртного Б. може бути представлено в наступному вигляді:
C6H12O6 + 2H3PO4 + 2АДФ ® 2CH3CH2OH + 2CO2 + 2АТФ.
Т. о., При зброджуванні 1 моля глюкози утворюються 2 благаючи етилового спирту, 2 благаючи CO2, а також в результаті фосфорилювання 2 молей АДФ утворюються 2 благаючи АТФ. Термодинамічні розрахунки показують, що при спиртному Б. перетворення 1 моля глюкози може супроводжуватися зменшенням вільної енергії приблизно на 210 кДж (50 000 кал), т. Е. енергія, акумульована в 1 молі етилового спирту, на 210 кДж (50 000 кал) менше енергії 1 моля глюкози. При утворенні 1 благаючи АТФ (макроергічних - багатих енергією фосфатних сполук) використовується 42 кдж (10 000 кал). Отже, значна частина енергії, що звільняється при спиртному Б., запасається у вигляді АТФ, що забезпечує різноманітні енергетичні потреби дріжджових клітин. Таке ж біологічне значення має процес Б. і у ін. Мікроорганізмів. При повному згорянні 1 моль глюкози (з утворенням CO2 і H2O) зміна вільної енергії досягає 2,87 МДж (686 000 кал). Інакше кажучи, дріжджова клітина використовує лише 7% енергії глюкози. Це показує малу ефективність анаеробних процесів в порівнянні з процесами, що йдуть в присутності кисню. При наявності кисню спиртове Б. пригнічується або припиняється і дріжджі отримують енергію для життєдіяльності в процесі дихання. Спостерігається тісний зв'язок між Б. і диханням мікроорганізмів, рослин і тварин. Ферменти, що беруть участь в спиртному Б., є також в тканинах тварин і рослин. У багатьох випадках перші етапи розщеплення цукрів, аж до утворення піровиноградної кислоти, - загальні для Б. і дихання. Більше значення процес анаеробного розпаду глюкози має і при скороченні м'язів (див. гліколіз ), Перші етапи цього процесу також схожі з початковими реакціями спиртного Б.
Зброджування вуглеводів (глюкози, ферментативного гідролізату крохмалю, кислотних гідролізатів деревини) використовується в багатьох галузях промисловості: для отримання етилового спирту, гліцерину і ін. Технічних і харчових продуктів. На спиртному Б. засновані приготування тесту в хлібопекарській промисловості, виноробство і пивоваріння.
Молочнокисле Б. Молочнокислі бактерії підрозділяють на 2 групи - гомоферментативное і гетероферментативні. Гомоферментативное бактерії (наприклад, Lactobacillus delbr ü ckii) розщеплюють моносахариди з утворенням двох молекул молочної кислоти відповідно до сумарного рівнянням:
C6H12O6 = 2CH3CHOH · COOH.
Гетероферментативні бактерії (наприклад, Bacterium lactis aerogenes) ведуть зброджування з утворенням молочної кислоти, оцтової кислоти, етилового спирту і CO2, а також утворюють невелику кількість ароматичних. речовин - диацетила, ефірів і т.д.
При молочнокислом Б. перетворення вуглеводів, особливо на перших етапах, близько до реакцій спиртного Б., за винятком декарбоксилювання піровиноградної кислоти, яка відновлюється до молочної кислоти за рахунок водню, одержуваного від НАД-Н. Гомоферментативное молочнокисле Б. використовується для отримання молочної кислоти, при виготовленні різних кислих молочних продуктів, хліба і в силосуванні кормів в сільському господарстві. Гетероферментативних молочнокисле Б. відбувається при консервуванні різних плодів і овочів шляхом квашення.
Маслянокислое Б. Сбраживание вуглеводів з переважним утворенням масляної кислоти виробляють багато анаеробні бактерії, що відносяться до роду Clostridium. Перші етапи розщеплення вуглеводів при маслянокислом Б. аналогічні соответстветственним етапах спиртного Б., аж до утворення піровиноградної кислоти, з якої при маслянокислом Б. утворюється ацетил-кофермент A (CH3CO-KoA). Ацетил-KoA може служити попередником масляної кислоти, піддаючись наступним перетворенням:
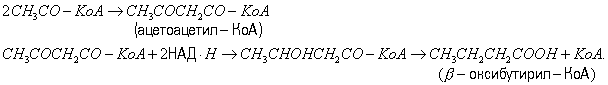
Маслянокислое Б. застосовувалося для отримання масляної кислоти з крохмалю.
Ацетоно-бутилове Б. бактерії Clostridium acetobutylicum зброджують вуглеводи з переважно. освітою бутилового спирту (CH3CH2CH2CH2OH) і ацетону (CH3COCH3). При цьому утворюються також в порівняно невеликих кількостях водень, CO2, оцтова, масляна кислоти, етиловий спирт. Перші етапи розщеплення вуглеводів ті ж, що і при спиртному Б. Бутиловий спирт утворюється шляхом відновлення масляної кислоти:
CH3CH2CH2COOH + 4H = CH3CH2CH2CH2OH + H2O.
Ацетон же утворюється декарбоксилюванням ацетоуксусной кислоти, яка виходить в результаті конденсації двох молекул оцтової кислоти. Дослідженнями В. Н. Шапошникова показано, що ацетоно-бутилове Б. (як і ряд ін., Наприклад пропіоновокисле, маслянокислое) в дослідах зі зростаючою культурою відбувається в дві фази. В першу фазу Б. паралельно з наростанням біомаси накопичуються оцтова і масляна кислоти; в другу фазу утворюються переважно ацетон і бутиловий спирт. При ацетоно-бутиловом Б. зброджуються моносахариди, дисахариди і полісахариди - крохмаль, інсулін, але не зброджуються клітковина і геміцелюлоза. Ацетоно-бутилове Б. використовувалося для промислового отримання бутилового спирту і ацетону, що застосовуються в хімічній і лакофарбовій промисловості (див. Також Ацетоно-бутилове бродіння і Ацетоно-етилове бродіння ).
Зброджування білків. Деякі бактерії з роду Clostridium - гнильні анаероби - здатні зброджувати не тільки вуглеводи, а й амінокислоти. Ці бактерії більш пристосовані до використання білків, що розщеплюються ними за допомогою протеолітичних ферментів до амінокислот, які потім піддаються Б. Процес зброджування білків має значення в круговороті речовин в природі (див. гниття ).
Пропіоновокисле Б. Основні продукти пропіоновокислого Б., що викликається декількома видами бактерій з роду Propionibacterium, - пропіонова (CH3CH2OH) і оцтова кислоти і CO2. Хімізм пропіоновокислого Б. сильно змінюється в залежності від умов. Це, мабуть, пояснюється здатністю пропіонових бактерій перебудовувати обмін речовин, наприклад залежно від аерації. При доступі кисню вони ведуть окислювальний процес, а в його відсутності розщеплюють гексози шляхом Б. Пропіонова бактерії здатні фіксувати CO2, при цьому з піровиноградної к-ти і CO2 утворюється щавелевоуксусная к-та, що перетворюється на янтарну к-ту, з якої декарбоксилюванням утворюється пропіонова к-та:
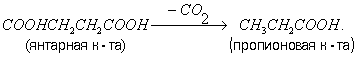
Існують Б., які супроводжуються і відновними процесами. Прикладом такого «окислювального» Б. служить лимоннокисле Б. Багато цвілеві гриби зброджують цукру з утворенням лимонної кислоти. Найбільш активні штами Aspergillus niger перетворюють до 90% спожитого цукру в лимонну кислоту. Значна частина лимонної кислоти, використовуваної в харчовій промисловості, виробляється мікробіологічним шляхом - глибинним і поверхневим культивуванням цвілевих грибів.
Іноді за традицією і чисто окислювальні процеси, здійснювані мікроорганізмами, називається Б. Прикладами таких процесів можуть служити уксуснокислое і глюконовокіслое Б.
Оцтовокисле Б. Бактерії, що відносяться до роду Acetobacter, окислюють етиловий спирт в оцтову кислоту відповідно до сумарної реакцією:
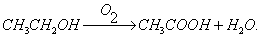
Проміжне з'єднання при окисленні спирту в оцтову кислоту - оцтовий альдегід. Багато оцтовокислі бактерії, крім окислення спирту в оцтову кислоту, здійснюють окислення глюкози в глюконовую і кетоглюконовую кислоти.
Глюконовокіслое Б. здійснюють і деякі цвілеві гриби, здатні окислювати альдегідну групу глюкози, перетворюючи останню в глюконовую кислоту:

Кальцієва сіль глюконової кислоти служить хорошим джерелом кальцію для людей і тварин.
Літ .: Шапошников В. Н., Технічна мікробіологія, М., 1948; Прескот С., Дан С., Технічна мікробіологія, пер. з англ., М., 1952; Пастер Л., Избр. праці, пров. з франц., т. 1-2, М., 1960; Кретович В. Л., Основи біохімії рослин, 4 видавництва., М., 1964; Фробішер М., Основи мікробіології, пер. з англ., М., 1965; Фердман Д. Л., біохімія, М., 1966; Работнова І. Л., Загальна мікробіологія, М., 1966.
В. І. Любимов.

Схема спиртового бродіння.
