МОСКОВСЬКИЙ ПСИХОЛОГО-СОЦІАЛЬНИЙ ІНСТИТУТ
РЕФЕРАТ з психофізіології НА ТЕМУ:
ГАМКергіческіх теорія депресії у людини
студентка:
КондратьеваД.А.
Група:
22ПМ / 4-08
викладач:
Москва 2006
зміст
Біологічні закономірності депресії ...................... ............ .3
ГАМК, тривога і депресія ...................................................... 5
Висновок ........................................................................ .11
Література ........................................................................ ..14
Біологічні закономірності депресії.
В останні роки актуальними є дослідження, пов'язані зі стресом і визначенням його вкладу в патогенез невротичних і афективних розладів.
Виявлено, що при депресивних розладах спостерігаються супрессия Т-клітинного імунітету, зниження кількості натуральних кілерів, дерегуляція факторів неспецифічної резистентності, активація гуморальних факторів імунітету. Імунні порушення пов'язані зі зміною нейроендокринної регуляції, що виявляється підвищенням рівня кортизолу та активацією нейромедиаторного обміну. Різний характер імунобіологічного реагування співвідноситься з особливостями клінічного поліморфізму афективних розладів. Біполярні афективні розлади характеризуються найбільш значущими порушеннями. У той же час, характерною особливістю псіхонейроіммунних взаємодій при хронічних афективних розладах (дистимии) є висока активність параметрів нейромедиаторного обміну (серотоніну і дофаміну) при нормальній концентрації гормонів. Використання кластерного аналізу дозволило виявити вченим взаємозв'язку клінічних характеристик депресивних розладів з імунологічними, нейромедіаторної і гормональними показниками, що доводить залученість цих гомеостатических систем в механізми псіхонейроіммуномодуляціі і в патогенез депресивних розладів. Ступінь вираженості депресивної симптоматики пов'язана з параметрами нейромедиаторного обміну, концентрацією адренокортикотропного гормону, кількістю хелперів / індукторів, а тривалість захворювання корелює з кількістю цитолитических кілерів / супресорів, натуральних кілерів, В-лімфоцитів.
Таким чином, основні закономірності псіхонейроіммуномодуляціі при депресії проявляються дисбалансом імунної, нейромедіаторної і гормональної систем.
Розглядаючи загальні нейрохимические, фармакологічні та нейрофізіологічні аспекти тривоги і депресії, вчені формулюють концепцію про взаємозв'язок тривоги і депресії в єдиний патогенний процес.
Стрес є провідним фактором патогенезу ЦНС, в першу чергу тривоги і депресії - найбільш поширених захворювань психіки [35, 50,51,52]. Гетерогенність механізмів патогенезу і форм тривоги і депресії, високий відсоток коморбидности, а також схожість багатьох клінічних проявів істотно ускладнює діагностику і терапію даних патологій [22,52]. За останні десятиліття висунуто безліч гіпотез про механізми виникнення тривоги і депресії [35,72], проте повна картина їх патогенезу ще не відома. Наявність однієї із зазначених патологій ускладнює терапію іншої патології, є предиктором поганого терапевтичного результату і фактором ризику виникнення інших захворювань ЦНС [50,51]. Це, а також схожість клінічних симптомів тривоги і депресії, їх генетичних особливостей і загальна залежність патогенезу від стресу вказують на те, що дані патології взаємодіють один з одним, а при виникненні тривоги і депресії можуть бути задіяні загальні механізми, біологічні маркери і фактори ризику [ 52].
Аналіз клінічної ефективності ряду антидепресантів (АТ) і анксіолітиків (АЛ) також підтверджуючого взаємозв'язок тривоги і депресії, оскільки синергетичний терапевтичний ефект при монотерапії надають багато препаратів обох класів, а поєднання АД і АЛ часто надає ефект, що перевершує суму ефектів кожного з них окремо [ 52]. На можливу спільність генезу тривоги і депресії вказують дані з області еволюційної психіатрії [47]. Важливим аргументом на користь спільності тривожно-депресивного патогенезу служать великі відомості з області експериментальної фармакології тривоги і депресії, які будуть розглянуті нижче. Таким чином, в сучасній біологічної психіатрії необхідні нові підходи до розуміння загальних механізмів виникнення тривоги і депресії.
Гамма-аміномасляна кислота (ГАМК) - найважливіший гальмові медіатор мозку, задіяний у багатьох процесах ЦНС, включаючи регуляцію емоцій, поведінки, пам'яті і вищих когнітивних функцій [35,46]. Існуючі клінічні та експериментальні дані дозволяють припустити, що саме ГАМК може бути тією сполучною ланкою, порушення якого призводить до патогенезу як тривоги, так і депресії, їх коморбідності, а також пояснює схожість симптомів і чутливість до одних і тих же препаратів.
ГАМК, тривога і депресія.
Протягом десятиліть в психофармакологии домінували амінергіческіе теорії патогенезу депресії, а патогенез тривоги традиційно пояснювався дефіцитом гальмівних медіаторів - серотоніну і ГАМК [2,3]. ГАМК-А рецептор являє собою складний пентамерний комплекс з безліччю сайтів зв'язування позитивних і негативних модуляторів, включаючи сайти зв'язування ГАМК, бензодіазепінів, стероїдів, алкоголю, барбітуратів, а також блокаторів хлорного іонофори [39]. Роль ГАМК і ГАМК-А рецепторів в патогенезі тривоги у людини і тварин добре доведена і детально обговорюється в цілому ряді робіт [2,35,51]. В цілому, придушення ГАМКергіческой фукнции призводить до тривоги, а активація центральних ГАМК-А рецепторів викликає анксіолізіс [35,50].
Набагато менш відома роль центральної ГАМК у патогенезі депресії, хоча наявні дані вказують, що участь ГАМК в механізмах виникнення депресії дуже важливо. Вперше ГАМКергіческіх гіпотеза депресії була сформульована в 1980 році [19] і за останні десятиліття набула значного підтвердження в клініці [11,45,67]. У таблиці 1 підсумовані клінічні та експериментальні дані про вплив депресії на ГАМКергіческіх систему мозку: в цілому, у людини і тварин спостерігається загальне зниження ГАМКергіческой функції при депресії і її відновлення під впливом різних антідепрессантних факторів [54,67,74]. Таким чином, з'являються підстави вважати, що саме ГАМКергіческіх система може участовать як в патогенезі тривоги і депресії [12,41], так і інтеграції даних патологій в єдиний тривожно-депресивний континуум (Рис. 1).
Малюнок 1. Роль ГАМК в інтеграції тривожно-депресивної патології (гіпотеза)



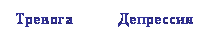


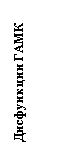

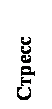







Грунтуючись на даних таблиці 1, слід очікувати, що препарати, що стимулюють ГАМКергіческіх систему, будуть ефективні і при терапії депресії. У таблиці 2 наводяться відомості за деякими ГАМК-активним АЛ, які в клініці дійсно виявляють виражене антидепресантну дію.
Таблиця. 1. Вплив депресії і антидепресантів на ГАМКергіческіх систему Клінічні дані Експериментальні дані При депресії · Зниження рівня ГАМК у плазмі, спинномозковій рідині і мозку у депресивних хворих [12,42,71] · Зниження рівня ГАМК у щурів в гіпокампі в моделі вивченої безпорадності [54 , 67], а також в корі і аккумбенсе в моделі вимушеного плавання - тесті Порсолта [11,26]. Підвищення щільності ГАМК-А рецепторів в септума у щурів в моделі вивченої безпорадності [40]. Підвищення зв'язування мусцімол у щурів, стійких в моделі поведінкового відчаю [16] · Зниження рівня ГАМК-активних стероїдів і підвищення рівня антагоністів стероїдів [27,37,38,46, 62,70], в тому числі у депресивних жінок з пре-менструальним синдромом [6] · Зниження рівня ГАМК-активних стероїдів (аллопрегнанолона) в корі в моделі соціальної ізоляції у мишей і щурів [6,62], дисбаланс рівня стероїдів в мозку в моделі ольфактобульбектоміі у щурів [76] · Зниження рівня пептиду-інгібітора зв'язування діазепаму в спинно-мозкової рідини і мозку у Депре ессівних хворих [61,65] і пацієнтів зі змішаною тривогою і депресією [29] · Зниження зв'язування бензодіазепінів в моделі хронічного стресу у фронтальній корі, гіпокампі, гіпоталамусі; підвищення рівня пептиду-інгібітора зв'язування діазепаму в мозку в моделі хронічного стресу у мишей [42] В результаті терапії не-ГАМКергіческіх антидепресантами · Підвищення рівня ГАМК в корі у депресивних пацієнтів [63] · Підвищення рівня ГАМК в гіпокампі у щурів в моделі вивченої безпорадності [ 40,54] · Підвищення рівня ГАМК в корі у пацієнтів після ЕСТ [64] · Підвищення рівня ГАМК у стріаутуме, мигдалині, гіпокампі і септума у щурів [26,31] Відновлення рівня ГАМК-активних стероїдних агоністів (аллопрегнанол вона) в плазмі у депресивних пацієнтів [37,38,76]; підвищення рівня ГАМК-активних стероїдних агоністів і придушення рівня антагоністів в плазмі у депресивних пацієнтів після одужання [62] · Підвищення рівня ГАМК-активних стероїдів (аллопрегнанолона) в мозку у щурів [62]
· Аналоги ГАМК і препарати, що впливають на метаболізм ГАМК:
Тіагабін [67], Бета-феніл-ГАМК [43], Габапентин [24, 28, 67], Амінооксіуксусная кислота [25, 53] (Також надає нормотіміческое дію [48]), Вігабатрін [8, 60]
· Бензідазепіновие агоністи:
Діазепам, карбамазепін і окскарбазепін [5, 50, 51, 53, 56]
· Етанол [37, 38]
· Стероїдні агоністи:
Прогестерон, тетрагідропрогестерон, аллопрегланолон [20] (Потенціюють дію інших АД [46]).
На підтвердження ГАМКергіческой теорії депресії у людини, Petty і Sherman [54] вперше вказали на роль ГАМК у патогенезі депресії у тварин. Згодом ці дані отримали подальше підтвердження в роботах цілого ряду лабораторій з використанням численних тварин моделей депресії [1,40,55]. Таблиця 2 підсумовує відомості про антидепресантною активності ГАМК-тропних АЛ при тестуванні в різних тварин моделях депресії. Доповнюючи загальну картину, в таблиці 3 наведені дані про продепрессантних властивості ГАМК-тропних анксіогенним препаратів у тварин.
Таблиця 2. антидепресантну ефекти ГАМК-активних анксиолитиков в експериментальних моделях депресії
препарати
Модель депресії і література
Аналоги препарати, що впливають на метаболізм ГАМК
вальпроат натрію
Амінооксіуксусная кислота
Вігабатрін
Гамма-вініл-ГАМК
Бета-феніл-ГАМК
· Вимушене плавання (тест Порсолта), миші [4,21,57]; потенціює ефекти інших АД в моделі вимушеного плавання, миші [73]
· Вимушене плавання, щури [10,11]
· Потенціює ефекти інших АД в моделі вимушеного плавання, миші [73]
· Ольфактобульбектомія, щури [36]
· Знімає викликане стресом підвищення щільності бензодіазепінових рецепторів в моделі вимушеного плавання, миші [58]
агоністи ГАМК
ГАМК
мусцімол
· Вимушене плавання, миші [4], вивчена безпорадність, щури [40,67]
· Вимушене плавання, миші, щура [4,10, 57]; потенціює ефекти АТ (етанолу і аллопрегнанолона) в моделі вимушеного плавання, миші [32,37,38]
Бензідазепіновие агоністи
діазепам
Карбамазепін та ін.
· Вимушене плавання, щури [49]; знижує муріцідную агресію в моделі ольфактобульбектоміі, щури [68]
· Вимушене плавання і вивчена безпорадність, миші, щура [7,34]; потенціюють інших АД в моделі вимушеного плавання, миші [73]
Етанол · Вимушене плавання, миші [32]; знижує депрессантное дію кокаїну в моделі вимушеного плавання, щури [30] Стероїдні агоністи
Прогестерон, тетрагидро-прогестерон
аллопрегнанолон
· Вимушене плавання, щури; потенціюють інших АД в моделі вимушеного плавання, миші, щура [20,62]
· Вимушене плавання, миші [37,62]; потенціює ефекти інших АД в моделі вимушеного плавання, миші [32]
Таблиця 3. Продепрессантние ефекти ГАМК-активних анксіогенним препаратів в експериментальних моделях депресії
препарати
Модель депресії і література Антагоністи ГАМК Бікукуллін · Вимушене плавання, вивчена безпорадність, миші, щура [4,30,54], знижує антідепрессантние ефекти вальпроату натрію, етанолу, фенгабіна і мелатоніну в моделях вимушеного плавання і ольфактобульбектоміі у щурів і мишей [21,32 , 44,57]
Бензодіазепінові інверсні агоністи
FG 7142, Ro 15-3505
Бета-карболіни-3-карбоксилату (БКК) етиловий та метиловий ефіри
· Вимушене плавання, миші, щура [13,69], вивчена безпорадність, щури [74]
· Вимушене плавання, щури [49], підвищена депресія в моделях вимушеного плавання і підвішування за хвіст у лінії мишей з природженою підвищеною чутливістю до метиловому ефіру БКК [17]
стероїдні антагоністи
Прегненолон-сульфат
· Вимушене плавання, миші [59]
Блокатори хлорного іонофори
Пікротоксин
коразол
· Вимушене плавання, щури та миші [4,69]; знижує антідепрессантние ефекти мусцімол, вальпроату натрію та інших АД в моделях вимушеного плавання і вивченої безпорадності у щурів [10,21,74]
· Вимушене плавання, щури [69]
В цілому, порівняльний аналіз даних, наведених в таблицях 1, 2 і 3, дозволяє зробити висновок, що у людини і тварин активації ГАМКергіческой системи відповідає зниження тривоги і депресії, тоді як придушення ГАМКергіческой активності корелює з підвищенням їх рівня. Все це підтверджує важливу роль ГАМКергіческой системи мозку в патогенезі тривоги і депресії, а також вказує, що пошук нових ГАМК-активних препаратів може бути перспективним напрямком при терапії тривоги і депресії.
висновок
Важливим аспектом в обговоренні ГАМКергіческой концепції тривоги і депресії є розгляд структур мозку, що беруть участь в патогенезі. Так, хронічний стрес - найбільш часта причина депресії - активує ГАМКергіческіе області переднього мозку, включаючи дорзомедіальний гіпоталамус і гіпокамп [31] - важливі елементи, традиційно відносяться до депрессогенних системі мозку. Численні роботи описують зміна морфології гіпокампу при депресії (див. [66]), вказуючи на його роль в патогенезі депресії. Гіпокамп багатий ГАМКергіческіх нейронами і є важливим компонентом мозкової ГАМКергіческой системи. Провідна роль гіпокампу в механізмах пам'яті, а також особлива роль негативних когниций в патогенезі тривоги і депресії, може бути одним з механізмів участі ГАМКергіческой системи мозку в патогенезі тривоги і депресії [35]. Морфологічні і метаболічні зміни при депресії були виявлені і в мигдалині [18,66] - ще однієї ГАМКергіческой структурі мозку, пов'язаної зі зберіганням і обробкою негативної пам'яті і одночасно бере участь в механізмах тривоги [15]. Підтверджуючи можливість патогенезу в рамках системи «тривога-депресія-пам'ять-мигдалина», Jasnow і Hubman [33] вказують на залучення ГАМКергіческіх процесів мигдалини в моделі депресії (обумовлене соціальне поразку) у гризунів. На важливу роль мигдалини в інтеграції тривоги і депресії вказують і клінічні дані з використанням функціонального магнітного резонансу [78], що відзначають виражену активацію мигдалини при пред'явленні зображень переляканих осіб депресивним пацієнтам. Добре також відомо про участь префронтальної кори в патогенезі різних типів тривоги [14,48]. При цьому префронтальная кора традиційно розглядається як частина «депресивної» патогенної системи мозку, морфологічні і метаболічні зміни при депресії в якій описані в багатьох роботах [66,78]. Відзначимо, що ГАМКергіческіе нейрони в ній складають найбільшу групу клітин. Крім того, префронтальна кора залучена в механізми пам'яті і вищі когнітивні функції, що може служити ще одним ГАМКергіческіх механізмом патогенезу в системі «тривога-депресія-каганець».
В цілому, наведені нами дані вказують на істотне перекривання ГАМКергіческіх структур мозку, залучених в патогенез тривоги і депресії. Це підтверджує наше припущення про існування в мозку загального ГАМКергіческіх патогенного кола при тривожно-депресивних розладах.
Як вже зазначалося раніше, генетичні фактори відіграють велику роль в патогенезі тривоги і депресії людини і тварин [23,48], і тому нейрогенетичному підходи до розуміння тривожно-депресивного патогенезу є надзвичайно перспективними. Переконливо демонструючи зв'язок ГАМК і депресії на нейрогенетичному рівні, в двох експериментальних моделях депресії (тесті Порсолта і підвішування за хвіст) генетичний локус, «відповідальний» за депресивний поведінку, було виявлено саме в області, що кодує одну з субодиниць ГАМК-А рецептора [77] (гени, що кодують субодиниці ГАМК-А рецепторів, важливі при патогенезі тривоги [75]).
Безумовно, кроме ГАМК в мозком існують и інші Механізми, что інтегрують патогенез тривоги и депресії. Так, Boyer [9] в статті «Чи мають тривога і депресія загальний патофізіологічний механізм?» Вказує на кортиколиберин в якості одного з таких можливих загальних патогенних ланок. Проте, на фармакологічному, нейрофизиологическом і нейрогенетичному рівні існують переконливі дані на користь того, що дисфункції ГАМКергіческой системи мозку, можливо, є провідним інтегральним патогенетичниммеханізмом тривоги і депресії. Тому пошук нових препаратів, що впливають на численні сайти ГАМКергіческіх рецепторів, може не тільки привести до появи нових ГАМК-активних АТ і АТ, а й сприяти створенню принципово нових типів психотропних засобів. Зокрема, ГАМКергіческіх препаратів, вибірково ефективних при терапії коморбидности; мнемотропних АЛ / АД-агентів, селективно пригнічують тривогу і депресію, пов'язані з негативними каганець; швидкодіючих ГАМК-активних АТ; препаратів для терапії змішаної тривоги і депресії (в тому числі неясною симптоматики і генезу); препаратів для лікування гострих нападів тривоги і депресії, в тому числі антипанічне і антисуїцидальних дії; АЛ і АТ, що вибірково діють в залежності від статі та ін. Пошук подальших терапевтичних впливів, спрямованих на загальний патогенетичний механізм, може дозволити на якісно іншому рівні вирішити проблему терапії тривоги і депресії.
література
1. І.В. Белозерцева, Б.В. Андрєєв, Ж-л ВНД, 47 (6), 1024 - тисяча тридцять один (1997).
2. А.В. Калу, Експ. клин. фарм., 60 (5), 3 - 7 (1997).
3. А.В. Калу, Н.А.Нуца, Експ. клин. фарм., 61 (5), 69 - 74 (1998).
4. KO Alley, SK Kulkarni, Naunyn Schmiedebergs Arch. Pharmacol., 339 (3), 306 - 311 (1989).
5. SV Argyropoulos, DJ Nutt, Eur. Neuropsychopharm., 9 (6), 407 - 412 (1999).
6. ML Barbaccia, S. Lello, T. Sidiropoulou et al., Psychoneuroendocrinol., 25, 659-675 (2000).
7. V. Benjamini, LL Skalitz, SRL Joka, R. Andreatini, Eur. J. Pharmacol., 347, 23 - 27 (1998).
8. FM Besag, Drug Saf., 24 (7), 513 - 536 (2001).
9. P. Boyer, Acta Psych Scand, 406, 24 - 29 (2000).
10. F. Borsini, S. Evangelista, A. Meli, Eur. J. Pharmacol., 121 (2), 265 - 268 (1986).
11. F. Borsini, A. Mancinelli, V. D'Arkano, S. Evangelista, A. Meli, Pharmacol. Biochem. Behav., 29 (9), 275 - 279 (1988).
12. L. Chang, CC Cloak, T. Ernst, J. Clin. Psych., 64 (3), 7 - 14 (2003).
13. K. Chopra, J. Kunchandy, SK Kulkarni, Arch. Int. Pharmacodyn. Thar., 294, 56 - 63 (1988).
14. RJ Davidson, Biol. Psych., 51, 68 - 80 (2002).
15. M. Davis, PJ Whalen, Mol. Psych., 6 (1), 13 - 34 (2001).
16. RC Drugan, SM Paul, JN Crawley, Brain Res., 631 (2), 270 - 276 (1993).
17. JC Do-Rego, C. Saudeau, G. Chapouthier, J. Costentin, Pharmacol. Biochem. Behav., 72, 411 - 416 (2002).
18. WC Drevets, Ann. NY Acad. Sci., 877, 614 - 637 (1999).
19. HM Emrich, DV Zervessen, W. Kissling et al., Arch. Psych. Nervenkr., 229, 1-16 (1980).
20. E. Estrada-Camarena, CM Contreras, M. Saavedra et al., Behav. Brain Res., 134, 175 - 183 (2002).
21. A. Fernandes Teruel, F. Boix, P. Escorihuela et al., Eur. J. Pharmacol., 152 (1-2), 1 - 7 (1988).
22. M. Freeman, SA Freeman, SL McElroy, J. Affect. Disorders, 68, 1 - 23 (2002).
23. DA Finn, MT Rutledge-Gorman, JC Crabbe, Neurogenet., 4, 109 - 135 (2003).
24. GCY Fong, BMY Cheung, CR Kurama, Med. Prog., 2, 1 - 8 (2003).
25. WS Gilmer, Expert Opin. Pharmacother., 2 (10), 1597 - 1608 (2001).
26. R. Gomez, CR Vargas, M. Wajner, HMT Barros, Brain Res., 968, 281 - 284 (2003).
27. MS George, A. Guidotti, D. Rubinov et al., Biol. Psych., 35 (10), 775 - 780 (1994).
28. SN Ghaemi, JJ Katzow, SP Desai, FK Goodwin, J. Clin. Psych., 59 (8), 426 - 429 (1998).
29. A. Guidotti, Neuropharmacol., 30 (12), 1425 - тисячу чотиреста тридцять три (1991).
30. T. Hayase, Y. Yamamoto, K. Yamamoto, BMC Pharmacol., 2, 1 - 9 (2002).
31. JP Herman, A. Renda, B. Bodie, Biol. Psych., 53, 166 - 174 (2003).
32. K. Hirani, RT Khisti., CT Chopde, Neuropharmacol., 43, 1339 - 1350 (2002).
33. AM Jasnow, KL Hubman, Brain Res., 920 (1-2), 142 - 150 (2001).
34. SRL Joka, LL Scalitz, V. Benjamini et al., Eur. Neuropsychopharmacol., 10, 223 - 228 (2003).
35. AV Kalueff, DJ Nutt, Depression Anxiety, 4, 100 - 110 (1996-1997).
36. JP Kelly, AS Wrynn, BE Leonard, Pharmacol. Ther., 74 (3), 299 - 316 (1997).
37. RT Khisti, SN Penland, MJ VanDoren et al, World J. Biol. Psych., 3, 87 - 95 (2000).
38. RT Khisti, CT Chopde, SP Jain, Pharmacol. Biochem. Behav., 67, 137 - 143 (2000).
39. ER Korpi, G. Grunder, H. Luddens, Prog. Neurobiol., 67, 113 - 159 (2003).
40. ML Kram, GL Kramer, M. Steciuk et al., Neurosci Res., 38, 193 - 198 (2000).
41. JH Krystal, G. Sanacora, H. Blumberg et al., Mol. Psych., 7, 71 - 80 (2002).
42. A. Kugaya, G. Sanacora, NPLG Verhoeff et al., Biol. Psych., 53, 1 - 8, (2003).
43. IP Lapin, CNS Drug Dev., 7 (4), 471 - 481 (2001).
44. KG Lloyd, B. Zibkovic, D. Sanger et al., J. Pharmacol. Exp. Ther., 241 (1), 245 - 250 (1987).
45. KG Lloyd, B. Zibkovic, B. Skatton et al., Prog. Neuropsychopharmacol. Biol. Psych., 13 (3-4), 341 - 351 (1989).
46. RB Lydiard, J. Clin. Psych., 64, 1 - 7 (2003).
47. R. Nesse, Neurosсi Biobehav. Revs, 23, 895 - 903 (1999).
48. EJ Nestler, E. Gould, H. Manji et al., Biol. Psych., 52, 503 - 528 (2002).
49. H. Nishimura, Y. Ida, A. Tsuda, M. Tanaka, Pharmacol. Biochem. Behav., 33 (1), 227 - 231 (1989).
50. D. Nutt, Understanding Mental Health, Octopus Ltd, Bristol (2001), 1 - 3.
51. D.Nutt, Eur. Neuropsychopharm., 10 (4), 433 - 437 (2001).
52. DJ Nutt, JC Ballender, D.Sheehan, HU Whittchen, Int. J. Neuropsychopharmacol., 5, 315 - 325 (2002).
53. G. Obrocea, RM Dunn, MA Frye et al., Biol. Psych., 51, 253 - 260 (2002).
54. F. Petty, AD Sherman, Pharmacol. Biochem. Behav., 18 (4), 649-50 (1981).
55. F. Petty, J. Affect. Disorders, 34, 275 - 281 (1995).
56. RM Post, TA Ketter, K. Denicoff et al., Psychopharmacol., 128 (2), 115 - 129 (1996).
57. V. Raghavendra, G. Kaur, SK Kulkarni, Eur. Neuropsychopharmacol., 10, 473 - 481 (2000).
58. D. Reddy, TINS, 24 (3), 103 - 106 (2001).
59. L. Rago, RA Kiivet, A. Adojaan et al., Pharmacol. Toxicol., 66 (1), 41 - 44 (1990).
60. HA Ring, R. Crellin, S. Kirker, EH Reynolds, J. Neurol. Neurosurg. Psych., 56 (8), 925 - 928 (1993).
61. A. Roy, Neuropharmacol., 30 (12), тисячі чотиреста сорок один - 1444 (1991).
62. R. Rupprecht, Psychoneuroendocrinol., 28, 139 - 168 (2003).
63. G. Sanacora, GF Mason, DL Rothman, JH Krystal, Am. J. Psych., 154 (4), 663 - 665 (2002).
64. G. Sanacora, GF Mason, DL Rothman et al., Am. J. Psych., 160 (3), 577 - 579 (2003).
65. JJ Sandford, SV Argyropoulos, DJ Nutt, Pharmacol. Ther., 88, 197 - 227 (2000).
66. Y. Sheline, Biol. Psych., 54, 338 - 352 (2003).
67. IS Shiah, LN Yatham., YC Gau, GB Baker, Prog. Neuropsychopharmacol. Biol. Psych., 27, 419 - 423 (1998).
68. S. Shibata, H. Nakanishi, S. Watanabe, S. Ueki, Pharmacol. Biochem. Behav., 21 (2), 225 - 230 (1984).
69. T. Skrebuhova, L. Alikmats, V. Matto, Meth. Find. Exp. Clin. Pharmacol., 21 (3), 173 - 178 (1999).
70. B. Spivak, R. Maayan, M. Kotler et al., Psychol. Med., 30 (5), 1227 - одна тисячу двісті тридцять одна (2000).
71. I. Sundman-Erikkson, P. Alard, J. Affect. Disorders, 71, 29 - 33 (2002).
72. I. Sundstrom Poromaa, S. Smith, M. Gulinello, Arch. Women Ment. Health, 6, 23 - 41 (2003).
73. G. Szymczyk, I. Zebrovska-Lupina, Pol. J. Pharmacol., 52 (5), 337 - 344 (2000).
74. G. Tunnicliff, E. Malatynska, Neurochem. Res., 28 (6), 965 - 976 (2003).
75. O.Yu. Vekovisheva, Academic Dissertation, Tampere (2003).
76. M. Uzunova, M. Ceci, C. Kohler et al., Brain Res., 976, 1 - 8 (2003).
77. T. Yoshikawa, A. Watanabe, Y. Ishitsuka et al., Genome Res., 4, 357 - 366 (2002).
78. DA Yurgelun-Todd, SA Gruber, G. Kanayama et al., Bipolar Disorders, 2 (3), 237 - 248 (2000).
79. Вісник біологічної психіатрії №12, 2003р. - електронний бюлетень РОБП і УОБП.
розділ: психологія
Кількість знаків з пробілами: 24378
Кількість таблиць: 3
Кількість збережений: 1
.... Почався пошук принципово нових рішень. 1.2 Ноотропил і інші лікарські засоби На початку 60-х років бельгійські фармакологи отримали з'єднання, яке представляло собою гамма-аміномасляна кислота, згорнуту в кільце і забезпечену деякими додатковими радикалами. Професор К. Джурджеа і його співробітники встановили, що ця сполука значно покращує пам'ять і полегшує ...
.... 6. Формування патологічної алгіческім системи (ПАС), що включає різні рівні центральної нервової системи і визначальною протягом десятиліть і характер всіх компонентів патологічного болю: больовий перцепції, страждання і больового поведінки. Хронічний біль як загальнопатологічних проблема Що ж являє собою хронічна (патологічна) біль з позицій загальної патології? У вченні про хвороби ...
"Накази", зворотні за своєю спрямованістю: наприклад, судинах і зіниці - звузитися, м'язах сечового міхура - скоротитися і так далі. Вегетативна нервова система дуже чутлива до емоційного впливу. Печаль, гнів, тривога, страх, апатія, статеве збудження - ці почуття викликають зміни функцій органів, що знаходяться під контролем вегетативної нервової системи. Наприклад, ...
... на мозковий кровотік, динаміку розвитку постішеміческіх феноменів і психоневрологічний статус залишається невивченим. Зазначені обставини і послужили підставою для порівняльного вивчення впливу селеніту натрію і селеніту цинку на системну гемодинаміку і мозковий кровотік в умовах норми і при патологічних станах. ГЛАВА 2 Матеріали та методи дослідження 2.1 Вивчення гострої ...
Так, Boyer [9] в статті «Чи мають тривога і депресія загальний патофізіологічний механізм?Хронічний біль як загальнопатологічних проблема Що ж являє собою хронічна (патологічна) біль з позицій загальної патології?