Хромосома людини в чотирьох вимірах
Н.Б. Рубцов
Микола Борисович Рубцов, д.б.н., зав. лаб. морфології і функції клітинних структур Ін-ту цитології і генетики СВ РАН (Новосибірськ)
У березні 2000 р на весь світ голосно і гордо прозвучало: "Геном людини секвенирован!". Однак скоро стало ясно, що геном людини прочитаний, але не розшифрований. Сьогодні доводиться визнати, що ми навчилися читати і писати, майже не розуміючи того, що пишемо і читаємо. Поки нам відомі лише деякі "слова" і зрозумілі деякі правила цього нового мови, вивчення якого, можливо, виявиться більш складним завданням, ніж просте "читання-записування" в довгі ланцюжки символів A, T, G, C. На цьому шляху нас явно чекають численні проблеми. Одна з них полягає в тому, що геном, записаний поки в одновимірному варіанті, насправді трьох- або навіть четирехмерен.
В даний час одне з важливих напрямків у вивченні реального генома - розшифровка просторової організації хромосом і їх окремих районів. Наші дослідження хромосомних перебудов показали, що реорганізація навіть однієї окремої хромосоми може змінити загальну архітектоніку ядра.
Хромосоми вчора і сьогодні
Історія вивчення хромосом цікава і повчальна. Звертаючись до неї, мимоволі згадуєш деякі з дитинства знайомі мудрі вислови і притчі. Наприклад, що шукати щось краще за все там, де світло. А історія обстеження слона групою сліпих показує, наскільки наші уявлення про об'єкт залежать від того, до якої його частини ми змогли дотягнутися. Довгий час вивчення хромосом ссавців в якійсь мірі сильно нагадувало цей процес. У більшості клітин хромосоми можна було побачити лише під час відносно короткого відрізка часу, пов'язаного з поділом клітини, тобто під час мітозу. Звичайно, є чимало винятків. У деяких клітинах, у деяких видів (майже як в деякому царстві, деякій державі) знайшли величезні хромосоми, видимі навіть неозброєним оком, причому під час інтерфази, коли в звичайних клітинах хромосома залишається невидимою і активно працює. Ці виняткові хромосоми виявилися конгломератом більше 1000 копій вихідної хромосоми. Найвідоміші приклади таких гігантів - політенія Drosophila melanogaster і Chironomus tummy.
Хромосоми ссавців, в тому числі і людини, набагато менше політенних. Однак труднощі їх аналізу полягала не в малому розмірі: для біологів, озброєних електронним мікроскопом, хромосома людини велика. Проблема в іншому. У звичайних клітинах в інтерфазі хромосоми так щільно прилягають один до одного, що навіть при самому дбайливому зверненні їх матеріал так перемішується, що не залишається жодних шансів розглянути окремі хромосоми, не кажучи вже про які-небудь деталі. Але незважаючи на всі ці труднощі, цитогенетики за допомогою електронної мікроскопії чимало зробили у вивченні організації хромосоми і її окремих елементів. Треба віддати належне і політенія, які довго розглядали як неперевершену модель інтерфазної хромосоми.
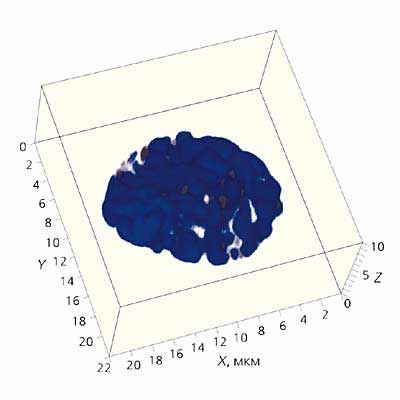
Мал.1. Тривимірна реконструкція хромосом фибробласта бурозубки в мітозі.
Синім кольором позначено ДНК хромосом, темно-сірим - теломерні райони;лініями - район сканування.
Масштаб вказано по осях X, Y і Z. Конфокальна мікроскопія проведена на мікроскопі LSM510META виробництва фірми ZEISS.
Повернемося до хромосом людини. Вони перетворюються в міцні структури лише на короткий час, безпосередньо перед поділом. Саме на стадії мітозу з'являється шанс розглянути їх досить докладно (рис.1). Правда, попередньо їх треба відокремити одну від іншої і розкласти на склі. Це завдання довго залишалося непосильним. З моменту перших описів хромосом в другій половині XIX ст. пройшло близько 80 років, перш ніж цитогенетики змогли правильно підрахувати число хромосом у людини. Це зробили в 1956 р практично одночасно дві пари дослідників: Дж.Тіо і А.Леван, Ч.Форд і Дж.Хаммертон. Їх роботи стали переломним моментом у вивченні хромосом людини.
В кінці 60-х - початку 70-х після розробки методів диференціального фарбування з'явилася можливість ідентифікувати всі хромосоми людини та їх окремі райони. Оскільки в таких дослідженнях хромосоми виглядали як витягнуті структури, що складаються з ділянок з різною інтенсивністю фарбування (їх шириною і товщиною можна було знехтувати), для їх опису досить було перерахувати порядок і орієнтацію всіх ділянок (рис.2).

Мал. 2. GTG-забарвлені метафазні хромосоми людини (47, XY, +18).
З огляду на, що основу хромосоми становить неймовірно довга молекула ДНК, яка в свою чергу описується як чергування пуринових і піримідинових основ (-ATGGTCACCTAGTTC-), ми звикли сприймати хромосому, з одного боку, як окремий елемент геному, з іншого - як одновимірну структуру. На цих уявленнях будувалася стратегія гібридизації для визначення локалізації конкретних генів спочатку з точністю до хромосоми, а потім до її району. Проводячи картування генів людини та інших ссавців, ми, як і інші дослідники, що працюють в цій галузі, стежили, спільно з якою хромосомою, а потім і її районом втрачається цікавий для нас ген [ 1 ].
Наші уявлення про хромосомі майже не змінилися, навіть коли з'явилися методи локалізації в хромосомі конкретних послідовностей ДНК за допомогою гібридизації нуклеїнових кислот in situ (безпосередньо на своєму місці). Цей метод багато в чому сприяв прочитання генома людини. Але в результаті подальших досліджень стало ясно, що даних про послідовності нуклеотидів в ДНК явно недостатньо, щоб реально розшифровки генома людини. Зусиллями молекулярних біологів і біоінформатики отримані деякі уявлення про тривимірної організації генома, але ці успіхи зазвичай обмежені порівняно невеликими його ділянками. Спроби провести реконструкцію тривимірної організації всього генома людини в віртуальному комп'ютерному світі (in silico) дали поки вельми обмежені результати. Можливо, біоінформатики і зможуть вирішити деякі питання, але перевірку знайдених ними відповідей все одно доведеться проводити експериментаторам.

Мал. 3. Часткова галерея оптичних зрізів інтерфазних ядра фібробластів людини.
Червоним виділена ДНК хромосоми 9; зеленим - ДНК центромерного району хромосоми 9; сірим - ДНК всіх хромосом.
Сьогодні прийшли нові технології і методи, завдяки чому мікроскопія в біології стала тривимірної. З'явилася можливість розглянути хромосому в інтерфазних ядрі і отримати інформацію про локалізацію в ньому відразу всіх хромосом людини [ 2 ]. Для цього широко застосовують гібридизацію in situ (FISH) ДНК індивідуальних хромосом, меченной флуоресцентними барвниками, з ДНК інтерфазних ядра. Потім за допомогою лазерного скануючого мікроскопа отримують серію оптичних зрізів ядра, на яких зареєстровані цікавлять дослідника сигнали (рис.3). Такі оптичні зрізи можна розглядати окремо, використовувати для створення ортогональних проекцій (рис.4) або для реконструкції тривимірної організації клітинного ядра (рис.5).
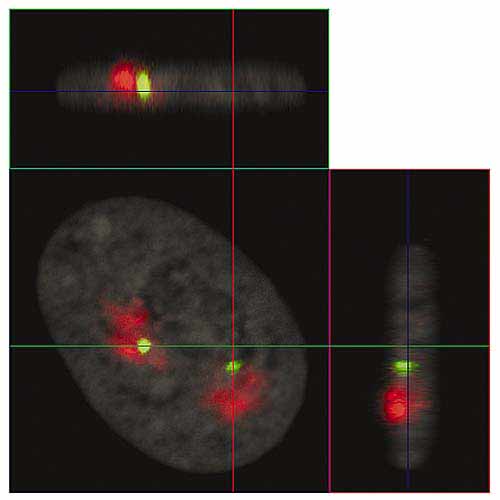
Мал. 4. Ортогональні проекції інтерфазних ядра фібробластів людини (серії їх оптичних зрізів наведені на рис.3).
Лінії показують положення площин оптичних зрізів: XY - синя, XZ - зелена, YZ - червона.
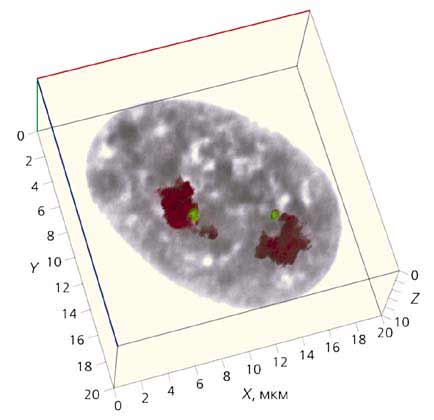
Мал. 5. Тривимірна реконструкція інтерфазних ядра фібробластів людини
із серії оптичних зрізів, наведених на рис.3.
Масштаб вказано по осях X, Y, Z.
Дивно, як мало часу було потрібно, щоб перейти до роботи в чотиривимірному просторі. Ще однією координатою стало час. Результати пошуку в інтернеті за словами 4D-microscopy (3D time-lapse microscopy) демонструють наочні докази прогресу в цьому напрямку. Те, що зовсім недавно здавалося абсолютно неможливим, сьогодні - передовий край досліджень, а завтра перетвориться, ймовірно, в рутину лабораторних буднів. Варто відзначити, що розроблені нами методи мікродіссекціі метафазних хромосом для отримання різноманітних ДНК-зондів внесли свій внесок у перетворення молекулярної цитогенетики в ефективну систему діагностики хромосомних аномалій.
Походження прекрасних метафазних хромосом
Акуратно розкладені на поверхні скла прометафазних і метафазні хромосоми ссавців радують око цитолога своєї чіткої морфологією вже більше 50 років. Коли милуєшся ними, не хочеться згадувати, що в живій клітині вони ніколи не бувають такими. Зі шкільної лави ми звикли до того, що в мітозі довжина хромосом зменшується. Малюнок на сторінках шкільного підручника наочно показує, як деконденсірованние в ядрі перед поділом хромосоми стають все коротшими і коротшими, перетворюючись в кінці кінців в компактні структури, які успішно розходяться в дочірні клітини (рис.6). За цими уявленнями варто багаторічний досвід тисяч цитогенетиків. Кожен день при аналізі цитологічних препаратів перед нашими очима проходили і сьогодні проходять тисячі подібних картин. У школі у цікавих дітей зазвичай виникало питання, як же відбувається така зміна розмірів і форми хромосом, і як вони розходяться, що не зав'язуючись в вузли?
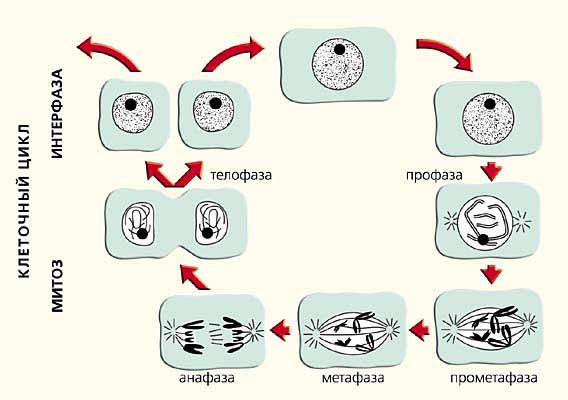
Мал. 6. Схема реорганізації хромосом у клітинному циклі (з шкільного підручника).
Тепер ми знаємо відповідь. Кожна хромосома займає в ядрі свою власну територію, що не ділячи її з іншими хромосомами. Виявилося, що не тільки хромосоми, але і їх плечі мають власні території. Наші спроби більш детального аналізу організації інтерфазних хромосоми показали, що окремі її райони розташовуються в займаному нею просторі приблизно в тому ж порядку, що і в метафазі. Більш того, розміри хромосоми в інтерфазі близькі до таких в метафазі. Висновок, який випливає з цих результатів, а також ряду інших робіт, в яких вдалося простежити за рухом конкретних ділянок хромосоми в ядрі від середини інтерфази до початку поділу, очевидний: при деконденсаціі-конденсації хромосом не відбувається жодних глобальних змін їх габаритів і значних переміщень хромосомних районів, тобто відношення довжини і ширини (товщини) хромосоми майже не змінюється.
Мимоволі виникає питання про походження довгих, красивих метафазних хромосом, які ми бачимо на цитологічному препараті. Завдяки їм можна ідентифікувати окремі хромосоми і їх райони, виявляти випадки внутрішньо-і міжхромосомні реорганізації, визначати локалізацію конкретних генів і інших послідовностей ДНК, а також робити багато інших корисних і потрібних речей. Але оскільки в живій клітині хромосоми за формою зовсім не схожі на метафазні, що лежать на предметному склі, то треба визнати, що такі хромосоми - просто артефакти. Для більшості цитогенетиків це дуже образливе слово. Хоча при спокійному міркуванні можна прийти до висновку, що засмучуватися нема чому. Так, в клітці не буває таких довгих хромосом, так, на цитологічному препараті вони виглядають так через спеціальної фіксації матеріалу і його особливого нанесення на скло. Що ж змінюється в хромосомі при переході клітини до мітозу, і чому ці зміни так впливають на морфологію хромосоми при укладанні на предметне скло?
Суть приготування препаратів митотических хромосом ссавців полягає в распластиваніе фіксованих клітин на поверхні скла. Цитоплазма розтягується до майже невидимого шару, а клітинні ядра перетворюються в плоский млинець. Найцікавіше відбувається з метафазної клітинами, в яких ядерна оболонка вже розібрана. Весь матеріал цих клітин розтікається тонким шаром, і тільки хромосоми в змозі чинити опір силам, розтягають клітинний матеріал по поверхні скла. Але і для них такий вплив не залишається непоміченим: їхні плечі розпрямляються, а хроматиди розходяться. Звичайно, після розбирання ядерної оболонки відбувається деяке пересування хромосом і, ймовірно, невелика зміна їх розмірів. Але в метафазі, анафазе і телофазе положення хромосом залишається майже таким же, як у попередній интерфазе.
При входженні клітини в мітоз здатність хромосом зберігати свою форму і розмір збільшується за рахунок їх зв'язування з білками конденсінамі. В результаті змінюється укладання хроматину, і хромосоми стають більш стійкими до розтягування і распластиваніе на склі під час випаровування фіксатора. Крім цього, в мітозі з хромосомних плечей йде білок когезін, який утримує поруч сестринські молекули ДНК, що виникли після подвоєння вихідної ДНК. Вивільнення когезіна призводить до втрати зв'язку між сестринськими хроматидами, які при распластиваніе хромосом розходяться. В результаті двуплечего хромосоми утворюють класичні Х-подібні фігури, зображення яких вже багато років вважається символом хромосом.
Дещо складніше справа йде з районами хромосомних плечей, з якими білки конденсіни зв'язуються по-різному. В першу чергу здатність чинити опір розтягуванню набувають райони, збіднені генами (так звані G-бенди). За цією ознакою розрізняються і райони, збагачені активно працюють генами (R-бенду). Більш того, ймовірно, існує досить строгий порядок, згідно з яким конденсіни зв'язуються з різними R-бенду, визначаючи зміни мітотичної хромосоми від профази до метафази. Чим далі пройшла клітина в мітозі, тим менше залишається районів, здатних до розтягування, і тим коротше буде хромосома на цитологічному препараті.
Правда, є й інший момент в приготуванні цитологічних препаратів. Він пов'язаний з силою, що діє на хромосоми при їх распластиваніе. Сила ця залежить від комбінації декількох факторів, зміна яких складно помітити (наприклад, вологості повітря). Іноді вже на наступний день якість приготованих цитологічних препаратів катастрофічно погіршується, і відбувається це не з вини лаборанта. Механізм, відповідальний за розтягнення-распластиваніе хромосоми на склі, досить простий, і зацікавлений читач може знайти його опис в опублікованих статтях [ 2 ].
Хромосоми in vivo, in vitro і in silico
Особливо цікаво порівняння складу, локалізації та порядку конденсації окремих районів хромосом. В першу чергу реплицируются (а конденсуються в останню) райони, збагачені активно працюють генами. Їх розтягнення, ймовірно, - одна з основних причин виявлення в хромосомах чергуються районів, що відрізняються за інтенсивністю фарбування. У розкладеної на склі хромосомі виділяються райони R- і G-бендів.
Але реальна хромосома організована інакше. При вивченні розподілу районів, збагачених і збіднених генами (R- і G-бендів), з'ясувалося, що в інтерфазних хромосомі ці дільниці не чергується уздовж хромосоми, як на цитологічному препараті, а розподілені між внутрішньою частиною хромосомної території і її поверхнею. Ймовірно, таке розташування R- і G-бендів зберігається і в мітотичних хромосомах. Тільки в момент распластиванія хромосоми на склі більше пухкий і еластичний матеріал R-бендів в результаті розтягування хромосоми переміщається на позицію, яка співпадає з просторовою віссю хромосомних плечей (рис.7).
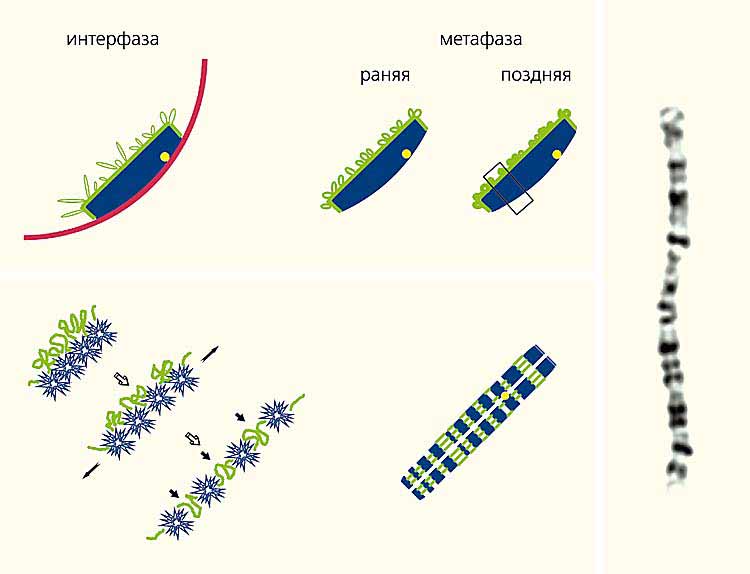
Мал. 7. Передбачувана схема реорганізації хромосоми в живій клітині в метафазну хромосому на цитологічному препараті.
На оптичних зрізах хромосоми (вгорі) видно райони, збіднені функціонуючімі генами (G-бенді, синій колір);збагачені активно Працюють генами (R-бенді, зелений);центромерного район (жовтий);ядерна оболонка (червоний).Внизу показано рух R- и G-бендів во время распластіванія хромосоми по склу (зліва) и їх Розташування в метафазної хромосомі на цітологічному препараті.Праворуч - Фотографія GTG-забарвленої другий хромосоми людини на цітологічному препараті.Показано, что в пространстве, займаною хромосоми, активно Працюють гени знаходяться на кордоні ее территории або в безпосередній блізькості від неї, а неактивних материал захованій Всередині ее территории. Ймовірно, це має величезне значення для регуляції актівності генів. Хромосомні територія Оточі межхроматіновім простором, в якому много ферментів, відповідальніх за транскріпцію генів и подалі превращение сінтезованої РНК [ 3 , 4 ]. Проникнуті у Внутрішній простір хромосомної территории ЦІМ ферментам непросто. Щільно упаковані хромосомний матеріал практично НЕ залішає вільного місця для їх пересування Всередині хромосомної территории. На Якийсь час Щільність упаковки ДНК зменшується. Без цього реплікація ДНК внутрішніх районів хромосоми би була Неможливо. Аджея ферменти реплікації повінні хоч на нетрівалій годину отріматі туди доступ. Разом з ними туди ма ють шанс проникнуті и ферменти транскріпції. Альо їх там НЕ может буті багато, и активно працювати там смороду не в змозі.
R- и G-бенді відрізняються НЕ только за кількістю и складом входять до них генів, щільності упаковки и просторової локалізації, а й за складом "нехромосомной" білків, что знаходяться в ціх районах. Порівняння ДНК R- і G-бендів показало чимало інших відзнак, наприклад за складом повторених послідовностей, по співвідношенню AT і GC пар нуклеотидів. Досить заглянути в будь-який підручник чи солідний огляд, щоб з'ясувати, що R-бенди збагачені GC парами і короткими дисперговані повторами, тоді як в G-бендах більше АТ пар і довгих диспергованих повторів.
Здавалося б, знання цих закономірностей дозволяє створити віртуальну модель хромосоми, та й усього генома людини. Однак така модель, побудована на основі відомої послідовності нуклеотидів з визначеними межами між R- і G-бенду, виявилася вельми далека від хромосоми in vitro (про хромосомі in vivo і говорити не варто). Але при порівнянні хромосоми in vitro з її віртуальною моделлю був вдало проведений пошук умов, при яких віртуальна модель виходила схожою на реальну хромосому. З'ясувалося, що збагачення AT або GC парами має значення, якщо порівнювати райони не в середньому по геному або окремої хромосомі, а по "найближчого оточення" (близько 9 · 10 6 пар нуклеотидів). У цьому випадку картина віртуальних хромосом виходила дуже близькою до схеми R- і G-бендів аналізованих хромосом.
Хромосома в n-мірному просторі
Пора згадати, що хромосоми - це об'єкти з реального світу, що займають в ядрі частина тривимірного простору. Якщо уявити час як ще одну вісь координат, то можна спробувати описати хромосому в термінах 4D. Як уже згадувалося вище, завдяки розвитку молекулярної біології і конфокальної мікроскопії можна не тільки "бачити" хромосому в клітинному ядрі (рис.5), але навіть відрізнити хромосомні території нормального гомолога від його перебудованого варіанту. Прикладом може служити наша робота по вивченню впливу реорганізації хромосом на їх становище в ядрах стовбурових клітин людини (рис.8).
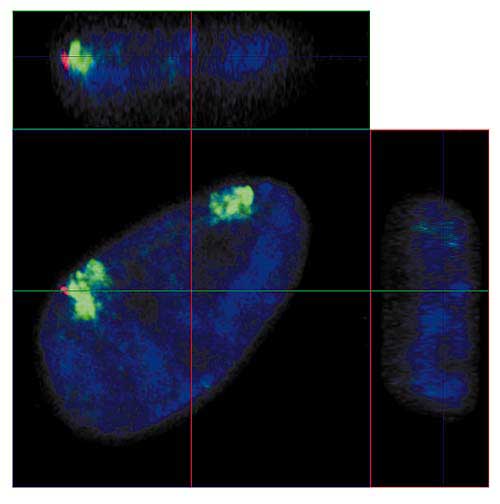
Мал.8. Ортогональні проекції інтерфазних ядра ембріональної стовбурової клітини людини,
в одному з гомологів хромосоми 18 якої втрачена частина короткого і частина довгого плеча.
Зеленим позначена ДНК хромосоми 18;червоним - ДНК з району делеции перебудованої хромосоми 18;синім - ДНК всіх хромосом.
Лінії показують положення площин оптичних зрізів як на рис.4.
На жаль, зазвичай життя складніше простих схем, а прості терміни можуть вносити значну плутанину. Сьогодні описати будова інтерфазних ядра можна без використання таких понять, як хромосомная територія і межхроматіновое простір [ 5 ]. Подивимося уважніше, що стоїть за цими термінами.
В даний час під "хромосомної територією" розуміють ту частину ядра, в якій за допомогою гібридизації міченої ДНК і тривимірної мікроскопії виявляється хромосомний матеріал. Межхроматіновим простором називають ту частину ядра, в якій при використанні стандартних методів і тривимірної мікроскопії хроматин не виявляється. Насправді частина матеріалу хромосоми знаходиться за межами її території, а межхроматіновое простір містить хроматин. У світлі цих даних широко використовувана термінологія виглядає дещо дивно. Але чи варто дивуватися? Ви впевнені, що коли-небудь бачили справжній білий колір? Чим він відрізняється від дуже світлого сірого? Чи не було там невеликого блакитного відтінку? Якщо немає проблем в розумінні того, що має на увазі ваш співрозмовник, використовуючи конкретний термін, то немає і проблем, пов'язаних з цим терміном.
Що ж являє собою матеріал хромосоми, наявний за її межами в межхроматіновом просторі? Розглянемо це на прикладі короткого плеча хромосоми 11 людини. Аналіз його локалізації в інтерфазних ядрі показав, що деякі ділянки молекули ДНК знаходяться далеко за межами хромосомної території. Цей факт легко пояснити. Окрема петля ДНК в ядрі, хоч я знаю при використанні хромосомоспеціфічних ДНК-зондів, в той час як FISH з ДНК-зондом, зробленим на базі клонованого фрагмента, дає в інтерфазних ядрі чіткий сигнал (рис.9).
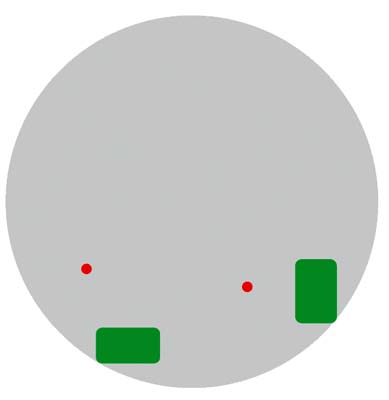
Мал. 9. ДНК хромосоми за межами хромосомної території (схема оптичного зрізу).
Сірим показано інтерфазна ядро; зеленим - територія короткого плеча хромосоми 11 [ 7 ].
Червоним - фрагмент ДНК з дистального району короткого плеча хромосоми 11.
Дійсно, з хромосомних територій в межхроматіновое простір виходять гігантські петлі ДНК. У їх складі присутні гени, які повинні активно працювати в клітці. Вони і йдуть за межі хромосомної території, тому що саме там умови для їх роботи ідеальні. У межхроматіновом просторі розташовані цілі комплекси молекул, що забезпечують синтез інформаційної РНК і її подальші перетворення. Є ще одна перевага для такого синтезу в межхроматіновом просторі. Воно не тільки місце локалізації фабрик синтезу РНК, а й магістральний шляхопровід, що дозволяє генним продуктам швидко дістатися до оболонки ядра, а потім через ядерну пору перейти в цитоплазму для участі у виробництві білка.
Зупинимося дещо докладніше на роботі таких "фабрик транскрипції" (термін, запропонований П.Фрезером, наочно відображає роль цих макромолекулярних комплексів). Одна така "фабрика" може одночасно працювати з ДНК з різних районів однієї хромосоми або навіть з різних хромосом. Причому відстань між генами однієї хромосоми може бути більш 20 млн пар основ. Це наочно показано в дослідах, проведених Фрезером і його колегами (рис.10).
 Мал.10. Локалізація активних генів в "блукаючих гігантських петлях" ДНК і "транскрипційних фабрик" в межхроматіновом просторі (схема оптичного зрізу).Сірим виділено інтерфазна ядро еритроїдної клітини миші, фіолетовим - території хромосоми 7, синім - "транскрипційні фабрики" (іммуноокрашіваніе РНК-полімерази II);червоний і зелений кольори - активні гени Hbb (b-ланцюга гемоглобіну) і Eraf (еритроїдного фактора) [ 5 ].
Мал.10. Локалізація активних генів в "блукаючих гігантських петлях" ДНК і "транскрипційних фабрик" в межхроматіновом просторі (схема оптичного зрізу).Сірим виділено інтерфазна ядро еритроїдної клітини миші, фіолетовим - території хромосоми 7, синім - "транскрипційні фабрики" (іммуноокрашіваніе РНК-полімерази II);червоний і зелений кольори - активні гени Hbb (b-ланцюга гемоглобіну) і Eraf (еритроїдного фактора) [ 5 ].
Крім гігантських петель, що виходять в межхроматіновое простір, є й інші райони хромосом, матеріал яких розташований на периферії їх територій і являє собою відносно слабко конденсований хроматин. У них також містяться функціонально активні гени, принаймні мають шанс бути активними, так як вони доступні для полимераз. Звичайно, перебування в цих районах не забезпечує включення гена, воно лише робить його доступним для включення. Дуже спокусливою виглядає гіпотеза, згідно з якою при диференціюванні клітин формується необхідний варіант просторової організації хромосомних територій: він визначає спектр генів, які можуть бути допущені до роботи і чия активність управляється іншими, звичними для молекулярного біолога механізмами.
Можливо, для повного опису хромосоми навіть розгляд її в чотиривимірному просторі виявиться недостатнім. Іноді дуже важлива і її віддалена історія, що виявляється в импринтинге відповідних хромосомних районів. При ідентичних геномах (по нуклеотидних послідовностей) можлива поява добре помітних порушень у їх власників. Так, невелика делеция в проксимальному районі довгого плеча хромосоми 15 людини може ініціювати і синдром Прадо-Віллі, і синдром Енгельмана. Визначальним у формуванні патології служить шлях хромосоми в зиготу: чи проходить вона через сперматогенез або оогенез.
Як не сумно, але слід визнати, що і сьогодні навряд чи можна розраховувати на повний опис хромосоми: її організації, стану та можливих потенційних змін, принципово важливих для реалізації в клітці інформації, записаної в її геномі. Але - дорогу здолає той, хто йде.
Значення оптимальної системи управління зрозуміло будь-якій людині. З результатами її помилок ми постійно зустрічаємося в нашому повсякденному житті. Організм людини набагато складніше господарства будь-якого навіть самого великого міста, і наслідки помилок в роботі його генома бувають досить сумні. Вивчення взаємодій елементів геному в пробірці або їх моделювання в комп'ютері - необхідні етапи вивчення генома людини, але їх явно недостатньо. Багато що визначається безпосередньо в клітці з неймовірно складною просторовою організацією її генома, для розуміння якої вже є ефективні інструменти. Хочеться вірити, що це завдання вдасться вирішити набагато швидше, ніж це представляється можливим сьогодні.
Робота виконана за підтримки Російського фонду фундаментальних досліджень. Проект 05-04-48221.
література
1. Жданова Н.С. Нитка Аріадни в генетиці // Природа. 2007. №6. С.18-25.
2. Рубцов Н.Б. Методи роботи з хромосомами ссавців. Новосибірськ, 2006.
3. Cremer T., Cremer C. // Nat. Rev. Genet. 2001. V.2. P.292-301.
4. Cremer T., Kreth G., Koester H. et al. // Crit. Rev. Eukaryot Gene Expr. 2000. V.10. №2. P.179-212.
5. Chakalova L., Debrand E., Mitchell JA // Nat. Rev. Genet. 2005. V.6. P.669-677.
6. Meaburn KJ, Misteli T. // Nature. 2007. V.445. P.379-381.
7. Mahy NL et al. // J. Cell. Biol. 2002. V.159. №5. P.753-763.
У школі у цікавих дітей зазвичай виникало питання, як же відбувається така зміна розмірів і форми хромосом, і як вони розходяться, що не зав'язуючись в вузли?
Що ж змінюється в хромосомі при переході клітини до мітозу, і чому ці зміни так впливають на морфологію хромосоми при укладанні на предметне скло?
Але чи варто дивуватися?
Ви впевнені, що коли-небудь бачили справжній білий колір?
Чим він відрізняється від дуже світлого сірого?
Чи не було там невеликого блакитного відтінку?
Що ж являє собою матеріал хромосоми, наявний за її межами в межхроматіновом просторі?