© В.А. Отелліні Формування патологій головного мозку
в ембріональний період
В.А. Отелліні
Володимир Олександрович Отелліні, член-кореспондент РАМН, професор,
керівник відділу морфології Інституту експериментальної медицини РАМН,
завідувач лабораторією онтогенезу нервової системи Інституту фізіології ім.І.П.Павлова РАН.
З 90-х років минулого століття, оголошених ООН Десятиліттям вивчення головного мозку, зросла кількість фундаментальних і прикладних досліджень, які охоплюють усі рівні організації мозку - від клітинного до системного і организменного. Особливу увагу медики і біологи приділяють проблемам формування центральної нервової системи людини в ембріональний період, коли закладаються основні структурні та функціональні характеристики, властиві зрілому мозку. В цей же час можливі і різноманітні збої в реалізації генетичної програми розвитку організму, які позначаються на формуванні плоду, а після народження можуть проявитися численними каліцтвами і нервово-психічні розлади. Ініціація і розвиток цих процесів нерідко обумовлені спадкової (хромосомної і генної) патологією, але іноді пов'язані з несприятливим впливом зовнішніх і внутрішніх факторів (забрудненням навколишнього середовища, підвищенням радіаційного фону, психологічними перевантаженнями і стресами, шкідливими звичками батьків і т. Д.).
За даними неонатологів, з кінця 90-х років спостерігається постійне зростання кількості неврологічних порушень у дітей різних вікових груп, причому в 27-44% випадків нервово-психічні розлади обумовлені порушеннями внутрішньоутробного розвитку [ 1 ]. Очевидно, що науковий пошук причин виникнення цих відхилень повинен бути спрямований на вивчення клітинних і тканинних особливостей нервової системи ембріонів, плодів та новонароджених дітей. При цьому важливо зрозуміти роль внутрішньоутробних структурних змін нервової системи в подальшому нервово-психічному розвитку дитини.
В останні роки помітно зросла кількість експериментальних робіт, присвячених вивченню впливу несприятливих факторів (радіації, алкоголю, нікотину, наркотиків і т. Д.) На ті чи інші функції головного мозку тварин. В результаті встановлено відмінності в проявах дозозалежних ефектів в структурах і функціях головного мозку на різних термінах ембріонального розвитку. Однак реєструються відхилення ніяк не співвідносилися зі клітинними і тканинними процесами, характерними для кожної ембріональної структури мозку в строго визначені терміни внутрішньоутробного розвитку - так звані критичні періоди.
У понятті «критичний період» немає нічого трагічного; це етап нормального розвитку зачатків органів, коли вони особливо чутливі до дії як біологічно активних речовин, що запускають і регулюючих клітинні і тканинні процеси генетичної програми розвитку, так і несприятливих факторів середовища, які деформують цю програму і призводять до формування патології. В пренатальний період (т. Е. До народження) кожен зачаток проходить один або кілька таких етапів, біологічний сенс яких полягає в тому, що створюється матеріальний фундамент, визначається наступний етап розвитку. Роль високої чутливості ембріонів під час критичних періодів полягає в підвищенні сприйнятливості організму до дії чинників, що забезпечують реалізацію спадкових потенцій.
Значення критичних періодів в індивідуальному розвитку організмів експериментально підтвердив видатний вітчизняний ембріолог П.Г. Светлов, який отримав в 1960 р за свою фундаментальну розробку Державну премію СРСР. Светлов, який назвав онтогенез реактивним процесом, детально вивчив на гризунах два критичних періоди: перший, попередній прикріплення (імплантації) зародка до стінки матки, коли відзначена найбільш часта загибель зародків після різноманітних впливів, і другий (імплантаційний), коли формуються зачатки органів і коли несприятливі фактори можуть сприяти виникненню різних вад [ 2 ]. Ці основні висновки узагальнив учень П.Г. Свєтлова - В.Ф. Пучков (рис. 1). Зрозуміло, що загибель ембріонів, пороки їх розвитку і потворності викликані порушенням процесів формування тканин і органів, проте хотілося б знати конкретні механізми виникнення патологій, оскільки без цього не можна розробити способи корекції та профілактики вроджених дефектів.
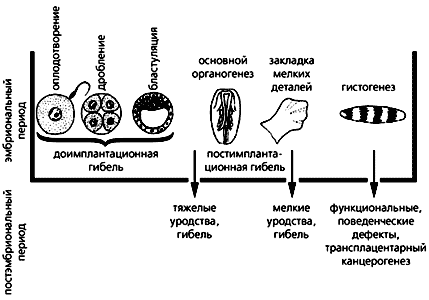
Рис.1. Дія факторів в критичні періоди ембріогенезу
і викликані цим наслідки. (За В.Ф. Пучкову.)
За спостереженнями клініцистів, відхилення в розвитку нервової системи найбільш часто пов'язані з перенесеною в пренатальний період гіпоксією (нестачею кисню), викликаної патологічним станом вагітної жінки або плода, забрудненням навколишнього середовища і т. Д. [ 3 ]. Гипоксические пошкодження ембріонального мозку після народження можуть сприяти розвитку епілепсії і церебрального паралічу, стати додатковим фактором ризику психічних і нейродегенеративних захворювань, вести до порушення поведінкових реакцій, а в особливо важких випадках - до загибелі дітей [ 4 ].
На жаль, більшість названих захворювань і розладів не піддається ефективному лікуванню, що в першу чергу пов'язано з відсутністю чітких уявлень про механізми і послідовності процесів (патогенезі), що призводять до розвитку конкретної форми хвороби. Без знання того, що, як і яким чином змінюється в головному мозку під впливом хвороботворних чинників, неможливо розробити стратегію лікування, спрямованого на корекцію порушених процесів, а не їх наслідків. Лікування, як вимагає сучасна медицина, має бути патогенетичним. Іншими словами, потрібно впливати на причини, що викликають розвиток хвороби, а не тільки на наслідки патологічних процесів. Саме тому необхідно експериментально вивчати формування структур головного мозку на тлі впливу несприятливих факторів середовища в ембріональний період, встановити характеристики, глибину змін різних відділів мозку в різні фази розвитку, що до сих пір практично не досліджено. Отримання таких даних складе основу для розробки нових, патогенетично обґрунтованих методів діагностики, лікування і профілактики вроджених захворювань центральної нервової системи.
У наших експериментах, в яких брали участь співробітники відділу морфології Інституту експериментальної медицини РАМН, лабораторії онтогенезу нервової системи Інституту фізіології ім. І.П. Павлова РАН та Інституту еволюційної біохімії і фізіології ім. І.М. Сеченова РАН, ми не обмежувалися анатомічним, макроскопічними рівнем. Вивчаючи ембріональний розвиток головного мозку ссавців (в тому числі і людини) при нормальному перебігу вагітності і на тлі впливів несприятливих факторів, ми використовували сучасні морфологічні (світлову і електронну мікроскопію, іммуноцітохіміі, нейротрансплантації, комп'ютерну морфометрію, поведінкові фізіологічні методи) і фізіологічні методи дослідження, що дозволяють оцінювати здатність до навчання і стан мотиваційно-емоційної сфери.
Нам треба було вирішити кілька завдань.
По-перше, необхідно було вивчити реакції функціонально різних формацій головного мозку народилися щурів (нової і старої кори мозку, базальних структур, стовбурових ядер - основних постачальників моноаминов мозку) на одноразові гіпоксичні впливу протягом пренатального періоду.
По-друге, - визначити залежність характеру змін в розвивається нервової тканини від стадії, на якій ембріони піддавалися гіпоксії.
Кора великих півкуль головного мозку була обрана в якості об'єкта дослідження далеко не випадково. Вона відрізняється чіткою впорядкованістю будови, а тому порівняно легко піддається якісному і кількісному аналізу відхилень у розвитку. У людини ця частина головного мозку досягла найвищого розвитку і бере участь в регуляції та побудові практично всіх функцій організму, включаючи пам'ять, пізнавальні і розумові процеси. Кора головного мозку поділяється на еволюційно давніші, старі (архикортекс) і нові (неокортекс) відділи, в яких виділяється близько 100 цитоархитектонических полів і підполів. Кожна ділянка відрізняється характерним будовою: своєрідним розподілом і співвідношенням типів нервових клітин (великих, малих і середніх пірамідних, зірчастих, веретеноподібних, біполярних) в шарах (яких в поперечнику кори може бути до семи-восьми), що розрізняються по товщині, щільності та орієнтації в них клітин. Всі ці клітини взаємопов'язані один з одним і з клітинами інших відділів мозку за допомогою систем асоціативних, проекційних і комісуральних шляхів. В межах цитоархитектонических полів або функціонально спеціалізованих зон кори мозку нервові клітини об'єднуються в так звані колонки (за їх відкриття в 1981 р Р. Сперрі, Д. Хьюбел і Т. Візел отримали Нобелівську премію), в яких обробляються приходять і виходять імпульси. В даний час є всі підстави стверджувати, що така складна і в певній мірі уніфікована конструкція кори головного мозку забезпечує її численні функції.
Перші ж результати експериментів виявилися для нас несподіваними. Одноразове годинне утримання щурів на різних термінах вагітності (13-й і 16-й день) в середовищі з недоліком (7.5%) кисню привело як до важких порушень загального розвитку потомства, так і до суттєвих структурних змін в центральній нервовій системі. Зазначалося зменшення розмірів і ваги плодів, скорочення обсягу їх мозку, розширення шлуночків мозку. Дія гострої гіпоксії в ці періоди, коли в корі активно протікають базисні гістогенетичних процеси (поділ клітин, їх переміщення і дозрівання), призводить до витончення і недорозвинення її шарів, порушення орієнтації та темпів дозрівання (диференціювання) нервових і гліальних клітин. У більш пізній постімплантаціонних період (19-й день), коли рівень поділу клітин у мозку знижується і прискорюються процеси їх дозрівання, гіпоксія надає менш виражений ушкоджує ефект. У всякому разі, що сформувалися шари кори незначно відрізнялися від контрольних ..
Мікроскопічний аналіз показав, що у новонароджених, п'яти- і шестиденних щурят, які перенесли гостру гіпоксію на 13-й і 16-й дні ембріогенезу, всі верстви кори стоншені і мають меншу, ніж в контролі, кількість нейронів, в тому числі великих пірамідних клітин, що беруть участь в тонкій координації рухових актів. Крім того, нервові клітини виглядають незрілими і їх орієнтація часто порушена.
Дещо інші зміни в неокортексе спостерігалися у потомства щурів, які перенесли вплив гіпоксії на 19-й день ембріонального розвитку. Дослідження мозку щурів на 10-й день після народження показало, що виявлене в попередні терміни деякий витончення верхніх шарів кори до цього часу нівелюється. Правда, у всіх шарах зустрічаються окремі загиблі клітини (п'ять-шість в поле зору). Таким чином, впливу факторів в більш пізні терміни постімплантаціонних періоду залишають в структурах мозку не аномалії і потворності, видимі неозброєним оком, а мікроскопічні відхилення в загальній компоновці і деталях будови кори головного мозку, недорозвинення нервових клітин, порушення їх взаємозв'язків, які можуть проявитися в подальшому житті тварини.
В паралельно проводяться фізіологічних дослідженнях було показано, що гіпоксія на 16-ту добу ембріогенезу призводить до зміни поведінки щурів: самці на відкритому просторі стають активнішими, виявляють тривогу, а самки, навпаки, - пассивнее. Більш того, у таких тварин відбувається інверсія статевої поведінки: самки починають вести себе як інтактні самці, а самці уподібнюються самкам. Цікаво, що недолік кисню на 19-ту добу внутрішньоутробного розвитку викликав у щурів обох статей однакова зміна поведінки на відкритому просторі - рівень тривожності і у самок, і у самців знизився.
Сьогодні ми дуже мало знаємо про механізми, що запускають пристосувальні і патологічні реакції у відповідь на дії факторів. Зрозуміло, що зовнішні подразники (біль, температура, радіація, отруйні речовини, недолік кисню і т.п.) опосередковують свої дії через різноманітні механізми - рецепторні, молекулярні, що в результаті трансформується в нервові процеси, що організують ті чи інші відповідні реакції. Істотна роль в цьому належить серотонинергическим механізмам (синаптическим і несінаптіческім), які забезпечують міжклітинні взаємодії за допомогою серотоніну - біологічно активного моноаміну, медіатора нервової системи і гормону. В ембріональний період онтогенезу серотонинергические нейрони контролюють процеси розвитку нервової тканини і відділів мозку, а після народження беруть участь в регуляції мотиваційно-емоційних компонентів поведінки, процесів навчання і пам'яті, больової чутливості та т. Д. Серотонінергічна системі відводиться важлива роль і в формуванні нервово-психічних захворювань і розладів (шизофренії, маніакально-депресивного психозу, стану тривожності і депресивності, мігрені), проте більшість механізмів і структурних виявлю ний цих порушень до теперішнього часу вивчено недостатньо.
У серіях наших експериментів зі зниженням концентрації серотоніну в різні критичні терміни внутрішньоутробного розвитку нам вдалося підтвердити виявлення П.Г. Свєтловим загальні закономірності реакцій ембріональних тканин на впливу факторів. Блокада парахлорфенілаланіном (ПХФА) ключового ферменту синтезу серотоніну - тріптофангідроксілази - в доімплантаційна період (до впровадження зародка в стінку матки) часто призводить до гальмування аж до повної зупинки процесів розвитку зародків. Порушується процес формування структур, що забезпечують харчування їх елементів, в тому числі внутрішньої клітинної маси, що дає початок всім клітинам і тканинам ембріонів (рис.2).

Зниження рівня ендогенного серотоніну на ранніх постімплантаціонних стадіях активного органогенезу (коли змінюється тип харчування зародка і формується плацента) призводило до недорозвинення плодів, утворення мозкових і пупкових гриж, мікроофтальмії, ущелинах твердого піднебіння (рис. 3). В результаті мікроскопічного дослідження були виявлені інші патологічні зміни в центральній нервовій системі плодів: великі крововиливи в різних відділах мозку (в довгастому мозку, смугастому тілі і в судинних сплетеннях шлуночків).
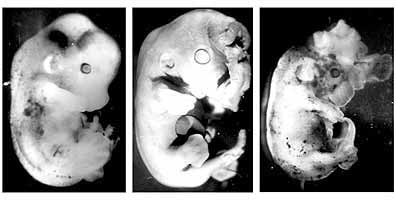
на восьмий день вагітності самок.
Видно мозкові грижі, підшкірні крововиливи.
Виникнення аномалій протягом ранніх стадій постімплантаціонних ембріогенезу може бути обумовлено виявленим нами зменшенням загального числа внутрімозкових серотонінергічних нейронів і їх аксонів після блокади синтезу серотоніну під час попередніх фаз розвитку. В результаті можливе виникнення вторинного дефіциту ендогенного серотоніну, який, як добре відомо, гальмує ділення (проліферацію) і дозрівання (диференціювання) нервових і гліальних клітин, що забезпечують всі функції головного мозку.
Виявилося, що в більш пізні стадії розвитку зародків, коли відбувається активне формування відділів мозку, зниження рівня ендогенного серотоніну призводить до порушення поділу клітин, зміни числа і напрямків їх міграції до генетично зумовленим ділянках мозку, що розвивається, а також затримується перетворення клітин-попередників (бластних форм ) в зрілі нервові і гліальні клітини. В результаті цього відбувається в більшій чи меншій мірі виражене недорозвинення одного або багатьох відділів мозку, формування аномалій або патологічних вогнищ.
Контроль (зліва) - у тварин, які розвивалися в звичайних умовах, впорядковані шари і строго вертикально орієнтовані відростки нейронів.
Досвід (праворуч) - у тварин, які розвивалися на тлі введення ПХФА, відбувається затримка дозрівання нервових клітин, збільшено їх число на одиницю площі, порушена орієнтація відростків нервових клітин.
Придушення синтезу серотоніну в критичний для неокортексу період (11-й день ембріогенезу) у мишей призводить до відхилень у розвитку і будові нової кори після народження: пригнічується ріст великих пірамідних клітин, при цьому їх кількість в V шарі зменшується, а в III і IV - збільшується (рис. 4). Крім того, підвищується кількість пірамідних клітин зі зміненою орієнтацією тел і верхівкових відростків дендритів. У нормі більшість таких дендритів незалежно від того, в якому шарі розташовується тіло клітини, спрямовані до поверхні кори майже перпендикулярно і досягають I шару, в якому утворюють своєрідне сплетіння. Саме ці апікальні дендрити є акцепторами всіх чутливих імпульсів, що приносять інформацію від органів чуття (зору, слуху, нюху і т. Д.), Серцево-судинної, травної, дихальної систем та опорно-рухового апарату. Перероблена в коркових нейронах інформація трансформується в виконавчі (еферентні) імпульси, що забезпечують адекватні сприйняття і регуляцію функцій організму. Складний, багатогранний і багатоступінчастий аналіз надходить в кору мозку інформації відбувається в кіркових вертикальних колонках клітин, які своєрідно і строго послідовно об'єднані між собою і з клітинами сусідніх колонок. Порушення ж міжнейронних взаємодій неминуче веде до різноманітних функціональних відхилень центральної нервової системи. В експерименті гіпоксія і дефіцит серотоніну в мозку ембріонів приводили до того, що понад 70% пірамідних клітин (в нормі їх близько 45%) відхилялися від вертикалі більш ніж на 45 °. На 5-й постнатальний день ці клітини концентрувалися в II-III і IV шарах, а на 10-й - в VI шарі. Ці дані виключно важливі, оскільки відображають участь серотоніну в порушенні ембріональної міграції клітин і в реорганізації міжнейронних зв'язків, які зберігаються в подальшому житті.
Після народження, в період до і на самому початку статевого дозрівання, у таких тварин відзначені відхилення в мотиваційно-емоційній сфері (знижувався рівень тривожності), в здатності до навчання (погіршувалися вироблення і відтворення умовного рефлексу пасивного уникнення). Причому, динаміка цих змін відрізнялася статевим диморфізму.
Таким чином, створюється враження, що дія різних факторів (будь то гіпоксія або дефіцит серотоніну, або ще щось) в певні критичні періоди ембріогенезу викликають подібні порушення в розвитку мозку, що призводять до формування Абнормальние структур, зміни клітинного складу і, ймовірно, міжнейронних зв'язків, що поєднується з відхиленнями в мотиваційно-емоційному поведінці після народження. Безумовно, багато що ще неясно, і поки рано говорити про медикаментозної корекції цих порушень. Дослідження необхідно продовжувати. Проте вже зараз необхідно залучити найширше увагу до профілактики трагічних наслідків нехтування здоровим способом життя і своєчасному лікуванню захворювань вагітних жінок, особливо в терміни від запліднення до завершення основного органогенезу у ембріонів (шість-вісім тижнів). Саме цей період найбільш небезпечний для життя і здоров'я потомства.
література
1. Володін М.М., Медведєв М.І., Рогаткин С.О. // Журн. невропатології і психіатрії ім.С.С.Корсакова. 2001. Т.101. №7. С.4-8.
2. Свєтлов П.Г. Фізіологія (механіка) розвитку. Л., 1978.
3. Neakas C., Buwalda B., Luiten P. // Progress in Neurobiology. 1996. V.49. №1. P.1-51.
4. Cannon T., Erg T.van, Rosso I. et al. // Arch. Gen. Psychiatry. 2002. V.59. №1. P.35-41.